

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Chronic hepatitis B virus (HBV) infection is considered as a major risk factor of chronic liver diseases including cirrhosis and hepatocellular carcinoma (HCC). Parenteral infection during the adulthood is a main route of chronic HBV infection in western countries, whereas vertical (or perinatal) transmission from mother to child is thought of a critical one in the East Asia (1, 2). Of importance, the modes of infection, together with other factors such as viral titer, genotype, genetic mutations of HBV, age and gender of the host may determine the long-term clinical course of chronic HBV infection. The mode of vertical transmission, in particular, is one of the most important determinants for the therapeutic responsiveness to antiviral therapies, or the natural course in terms of hepatitis e antigen seroconversion and even the development of complications (3, 4, 5).
Previous studies using a comparison of HBV DNA sequences and phylogenetic analyses have shown that HBV of chronically infected children originates mainly from their respective mothers or fathers (6, 7, 8). Although there has been hitherto a molecular evidence of vertical transmission of HBV in the pairs of mother-young child, it is improbable to identify exactly the mode of infection (especially vertical vs non-vertical) in real clinical settings because medical information from the adults for the source of HBV, relying on history taking alone, is highly obscure and there could be recall bias. There have been, so far, no reports about whether it was possible to demonstrate the molecular evidence of vertical transmission of HBV even after several decades of infection.
Taken together, 'Vertical or non-vertical transmission' is so crucially informative that it enables clinicians to manage effectively the patients with chronic HBV infection and also foresee the prognosis or therapeutic responses. Therefore, this study was conducted to prove the vertical transmission by showing the molecular closeness of HBVs between adult patients with clinically presumed vertical transmission and their respective mothers with the phylogenetic analyses of pre-S/S genes of HBV. It must provide valuable information to design a further strategy of management if the mode of HBV infection can be disclosed even after long-term period of infection.
MATERIALS AND METHODS
Subjects and definitions
A total of 36 subjects (18 mother-child pairs) were enrolled at Hallym University Medical Center, Seoul, Korea, and underwent clinical, biochemical, and virologic evaluations. Chronic HBV infection with vertical transmission was clinically defined on the basis of the following criteria; 1) patients were positive for hepatitis B surface antigen (HBsAg) during the whole lifetime, 2) patient's mothers were also positive for HBsAg, 3) patient's fathers were negative for HBsAg, 4) there was a significant clustering of HBsAg positive siblings, and 5) there were no other risk factors of the infection such as a history of blood transfusion, operation, parenteral drug abuses, needle stick injury, hemodialysis, or having multiple sexual partners. If the patients did not meet the criteria completely or there was no serum sample before an exposure to antiviral drugs, they were excluded from the study.
Serum samples were tested for HBsAg, antibody to HBsAg, hepatitis e antigen (HBeAg), antibody to HBeAg with chemiluminescent microparticle enzyme immunoassay (Abbott Laboratories, North Chicago, IL, USA). Serum HBV DNA levels were quantitated using the VERANT 3.0 assay (Bayer Healthcare, Tarrytown, NY, USA; lower limit of detection: 2,000 copies/mL) or COBAS TaqMan PCR assay (Roche, Branchburg, NJ, USA; lower limit of detection: 116 copies/mL).
The severity of chronic liver diseases was classified as follows: 1) inactive HBsAg carrier was defined as those who were positive to HBsAg for more than 6 months, negative to HBeAg, had a serum HBV DNA levels < 104 copies/mL and persistently normal serum alanine aminotransferase (ALT) levels for more than one year, 2) chronic hepatitis B as those who had a serum HBV DNA levels > 104 copies/mL, persistently or intermittently elevated serum ALT levels and a lack of evidence demonstrating cirrhosis and HCC (9), 3) cirrhosis was diagnosed by histology and/or ultrasonographic/CT imaging features, supplemented with clinically relevant evidence of portal hypertension (esophageal and/or gastric varices, ascites, splenomegaly with a platelet count of < 100,000/µL) or hepatic encephalopathy, 4) the diagnosis of HCC was made according to the guidelines of American Society for the Study of Liver Diseases, which was based on the imaging studies and/or serum alpha-fetoprotein levels (10).
HBV DNA extraction, amplication, and sequencing
HBV DNA was extracted from serum samples using commercial kits (QIAamp DNA Blood Mini Kit; Qiagen, Hilden, Germany) according to the manufacturer's protocol. Serum was collected and stored at -70℃ until use. Pre-S and S genes of the HBV DNA genome were amplified by nested PCR using the amplification primers and the thermocycling conditions described previously (11, 12). The sequencing primers for target regions are shown in Table 1. The predicted lengths of pre-S and S regions were 699 bp and 541 bp amplicon, respectively. These sequences covered the whole pre-S and partial S region.
The PCR products were then purified using a QIAEX II gel extraction kit (Qiagen, Hilden, Germany) and directly sequenced two times for each sample using ABI prism™ BigDye™ Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems Inc., Foster, CA, USA) on an ABI automated fluorescent sequencer (ABI 3730XL, Applied Biosystems Inc.).
HBV serotype and genotype
Since a previous study showed that most HBV serotypes in Korea were adr and adw, we determined the serotypes by analyzing amino acid sequences at positions 122 and 160 in the surface gene. The HBV genoypes were determined by phylogenetic analysis based on the partial sequences of S gene of the HBV DNA. Nucleotide sequences were compared with 32 reference sequences from the eight HBV genotypes (A-H) obtained from GenBank (Accession numbers of the representative subgenotypes in Fig. 1; A; AY233277, AF090841, AB194951, AM180623, B; AB010290, AB073830, M54923, AY033072, AB219429, C; AY217376, AB111946, Y18857, AY123041, X75656, X75665, AB048704, AB048705, AB241111, AY247030, AY247032, X14193, D; AB104711, AB205127, AY902770, AB033559, E; X75664, F; AY179735, X69798, AY311370, AF223965, G; AB056514, H; AY090454).
Phylogenetic analyses
HBV pre-S/S gene sequences were edited using BioEdit software version 7.0.9 (13). They were subjected to alignment with Clustal X software version 2.0 (14). Phylogenetic tree was constructed using the Neighbor-Joining algorithm implemented in the MEGA software version 4.0 (15) with the Kimura-two-parameter model of molecular evolution. The reliability of the inferred tree was assessed by a bootstrap resampling test including 1,000 replications.
RESULTS
Child group was 29.3 yr in mean age and male in 44.4% (8/18). Most patients were chronic hepatitis B status (17/18), only one patient has cirrhosis. Mother group, the mean age of 55 yr, had more frequently advanced liver diseases such as cirrhosis or HCC (61.1%, 11/18) than child group. The baseline demographic characteristics were listed in Table 2.
All isolates of HBV were genotype C and serotype adr, which is alleged as the most prevalent form in Korea (Fig. 1). The nucleotide homology of the preS and S regions among all the isolates of HBV was over 95%. The homology and divergence between mothers and children were 97.3%-99.8% and 0%-1.5%, respectively. Three patients (C13, M15, M17) had deletion mutations in the preS2 region ranging from 33 to 48 bp long. M15 and M17 patients had same deletion mutations. As for "a" determinant of HBsAg, there were four point mutations eliciting three amino acid substitutions in two sites. These did not elicit change of HBV subtype or serotype. These showed very similar patterns between child and mother, and summarized in Table 3.
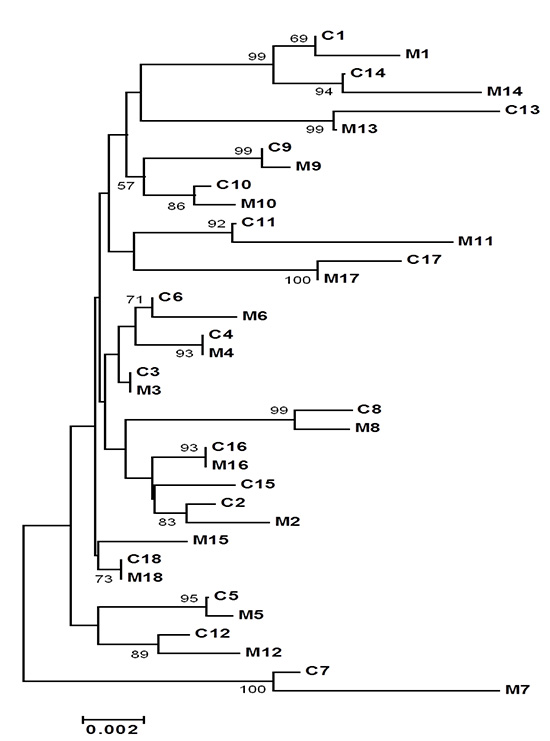
All families except Family 15 showed a great nucleotide homology to be more than 98%. Even though all HBVs showed great nucleotide homology, phylogenetic tree of HBV preS and S gene demonstrated that 17 out of 18 child-mother pairs (94%) were grouped into the same cluster (Fig. 2). Phylogenetic tree analysis of HBV preS gene showed same result of preS/S gene analysis (Fig. 3). Phylogenetic tree analysis of HBV S gene demonstrated also a similar tree topology (Fig. 4).
DISCUSSION
In the present study, HBVs from the adult patients with presumed vertical transmission were clustered together with HBVs from their respective mothers in 17 of 18 child-mother pairs (94%) when phylogenetic trees of pre-S/S genes of HBV were constructed. It strongly suggests that vertical transmission can be proven ever since several decades of the infection have elapsed.
Among DNA viruses, HBV is highly prone to mutations during evolution because of lacking proof-reading repair-enzyme activity of reverse transcriptase. Such a quasi species make HBV to be able to escape immune surveillance or vaccine. The most vulnerable part of genetic mutations in HBV is core promoter and pre-core region, followed by pre-S/S regions, though they may occur at any regions. It is of note that the mutations are related to viral loads and immune activity of the hosts, which may affect the clinical course of the patients (16). On the other hand, it is still controversial about what region of HBV genes is the most proper target for molecular analysis on the mode of transmission because of quasi species or frequent mutations. Accordingly, the selection of genetic targets among pre-S, S, or core/pre-core regions depends purely on the inclinations of investigators. Of HBV genes, S region is most conserved compared to core promoter and pre-core regions. It means that analysis of S region is, theoretically, most suitable toward the changes of HBV genes in long-term period while core promoter/pre-core region is rather appropriate for molecular analysis in short-term period such acute HBV infection or in the patients with a young age (6, 17, 18, 19, 20). In the present study, the age of patients with vertical transmission ranged from 19 to 55 yr, which means that the duration of HBV infection is from at least 19 to 55 yr at maximum. Therefore, core promoter/pre-core regions with a high potency of too excessive mutations could not be candidates on the study. Considering the quasi species of HBV, S gene, the most conserved region, was selected for phylogenetic analysis and then pre-S gene was analyzed in concert to supplement the findings. As described, both phylogenetic trees showed the similar topology. The most interesting finding is that it is possible to cluster the patients with vertical transmission and their respective mothers into the same group on phylogenetic trees, even in adult patients since long-term period, e.g., even several decades, of HBV infection has elapsed.
To date, some of HBV viral mutations, especially preS and S regions are known to be associated with disease progression (11, 22, 23, 24, 25). In the present study, preS deletion mutations were detected in 3 patients. Among them, two patients had advanced liver disease such as cirrhosis or HCC. It is compatible with previous study. When the "a" determinant is affected, important changes with regard to immunity and protection from HBV infection may arise. In our study, there were some point mutations in "a" determinant of S region eliciting amino acid substitutions. Although it did not show clinical significance in disease severity, mutations between mother and child revealed very similar patterns interestingly. It means viral evolution HBV in child followed by that in mother.
In the aspects of HBV genotype, all the subjects in this study had a genotype C, which is known to be most prone to mutations and also resistant to antiviral therapies such nucleot(s)ide analogues and interferon compared to the other genotypes (3, 21). Epidemiologically, not only vertical transmission but also genotype C is considered an influential risk factor for advanced liver diseases including cirrhosis and HCC. It is of importance to identify the routes or modes of HBV infection in scheming a therapeutic challenge and predicting the clinical prognosis, especially in the East Asia where vertical transmission is a main cause of chronic HBV infection. Both history taking on the clinic and laboratory findings, however, are not enough to tell whether HBV is vertically infected during the perinatal period (from mother) or adulthood (from others). If phylogenetic analyses of HBV pre-S or S genes were simply available in the future, therefore, patients with chronic HBV infection can be classified more delicately and managed more properly on the basis of vertical vs non-vertical transmission even in adult patients.
Our study had some limitations. We did not perform cloning to identify viral quasi species and analyze whole HBV genes including pre-core and core regions. Also, the subject number of this study was relatively small. Thus, a larger study including full HBV genes will be warranted in the future.
In conclusion, most pairs with presumed vertical transmission were grouped into the same cluster on phylogenetic trees of pre-S/S genes of HBV even after long-term period of infection elapsed. This study may suggest that vertical transmission can be proved even after several decades of HBV infection using phylogenetic analyses which may give a valuable clinical information to make a better strategy for chronic HBV infection.




 XML Download
XML Download