

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Ventilator-induced lung injury (VILI) refers to deleterious lung injury produced or worsened by mechanical ventilation (MV) (1). Acute respiratory distress syndrome (ARDS) patients receiving MV are at significant risk for developing VILI. Lung protective ventilation (LPV) with low tidal volume has been shown to reduce VILI and mortality in ARDS patients (2). However, the effectiveness of LPV may be limited because of the heterogeneity of lung involvement, resulting in an inability to completely prevent regional alveolar distension (3). To prevent VILI in effective, adjunctive therapeutic strategies based on a precise understanding of the pathophysiology of VILI need to be evaluated.
Transglutaminases (TGs) are enzymes that catalyze the post-translational modification of proteins through the formation of isopeptide bonds (4). They play a pivotal role in several biochemical processes such as blood clotting, skin formation, and apoptosis through the modification of various substrate proteins (5, 6, 7). Of the TGs, transglutaminase 2 (TG2) is considered a key factor in the protection from injury and in the promotion of repair (8). Aberrant induction of TG2 activity, however, contributes to various disease pathologies, including neurodegenerative diseases, atherosclerosis, inflammatory diseases, autoimmune diseases, and fibrosis (9). Increased TG2 activity is commonly detected in diseased tissues with inflammation and in cells undergoing inflammatory stress (10), and has been reported to induce or exacerbate inflammation via nuclear factor-κB (NF-κB) activation (10, 11). While several studies have explored the role of TG2 in lung disease, including in non-small cell lung cancer (12), pulmonary fibrosis (13), allergic asthma (14), and acute lung injury (15), this is the first study to assess the potential role of TG2 in VILI.
The purpose of this study was to examine: 1) the TG2 activity and gene expression in the inflammation which could be over-exaggerated by the injurious MV strategy, 2) the effect of TG2 inhibition on TG2 activity, gene expression, and inflammatory parameters, and 3) the additional effect of TG2 inhibition in the LPV strategy in a mouse VILI model.
MATERIALS AND METHODS
Animals and mechanical ventilation
Five-week-old specific-pathogen-free male C57BL/6 mice (OrientBio, Sungnam, Korea), each weighing 20-25 g, were randomly divided into the following six experimental groups: 1) a control group (n=24), in which mice were tracheostomized and instilled with 50 µL of saline; 2) a lipopolysaccharide group (LPS group, n=24), in which mice were instilled with 0.5 mg/kg of LPS (Escherichia coli O127:B8, Sigma, St. Louis, MO, USA) in 50 µL of saline through the tracheostomy; 3) a lung protective ventilation group (LPV group, n=24), in which mice were ventilated with low tidal volumes (VT) and positive end-expiratory pressure (PEEP) after instillation of 0.5 mg/kg of LPS in 50 µL of saline; 4) a ventilator-induced lung injury group (VILI group, n= 24), in which mice were ventilated with a high tidal volume without PEEP after instillation of 0.5 mg/kg of LPS in 50 µL of saline; 5) a VILI with cystamine pretreatment group (Cyst+VILI group, n=24), in which mice were pretreated with the TG2 inhibitor cystamine and ventilated with the same settings as in the VILI group after LPS instillation; and 6) a LPV with cystamine pretreatment group (Cyst+LPV group, n=24), in which mice were pretreated with the TG2 inhibitor cystamine and ventilated with the same settings as in the LPV group after LPS instillation. Each group was subdivided into four experimental subgroups: 1) a tissue subgroup (n=6) for histopathologic examination; 2) a bronchoalveolar lavage (BAL) subgroup (n=6) for measurement of tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 in BAL fluid (BALF); 3) a tissue homogenate subgroup (n=6) for measurement of NF-κB activity in lung tissue homogenates; and 4) a TG2 subgroup (n=6) for TG2 quantitative real-time polymerase chain reaction (RT-PCR) analysis and measurement of TG2 activity.
Tracheostomy and intubation were performed under anesthesia with an intraperitoneal injection of 65 mg/kg of pentobarbital sodium. MV was performed using a rodent ventilator (Harvard Apparatus, Holliston, MA, USA). The mice in the LPV group were ventilated with 7 mL/kg tidal volume, a PEEP of 3 cmH2O, and a respiratory rate of 90 breaths/min for 4 hr. An adequate setting for the VILI model with increase in TG2 activity has been determined by preliminary studies using various MV settings (16, 17, 18, 19). The mice in the VILI group were ventilated with 35 mL/kg tidal volume, a PEEP of 0 cmH2O, and a respiratory rate of 90 breaths/min for 4 hr. To maintain deep anesthesia, half of the initial dose of pentobarbital sodium was administered once every 1 hr of MV.
Evaluation of ventilator-induced lung injury
After MV, mice from the tissue, tissue homogenate, and TG2 subgroups were rapidly exsanguinated by dissection of the abdominal aorta. The heart and lungs were excised en bloc through a midsternal incision. The lungs of the tissue subgroup were immediately instilled with 4% paraformaldehyde through the trachea at a hydrostatic pressure of 15 cmH2O and fixed in 4% paraformaldehyde for 48 hr. Paraffin blocks were prepared by dehydrating samples with ethanol and embedding in paraffin. The posterior portions of the right lower lobe were sectioned at a thickness of 5 µm, placed on glass slides, and stained with hematoxylin-eosin (H-E). A pathologist blinded to the protocol and the experimental groupings examined the degree of lung injury and graded the specimens by acute lung injury (ALI) score based on: 1) alveolar capillary congestion; 2) hemorrhage; 3) infiltration or aggregation of neutrophils in the airspace or the vessel wall; and 4) the thickness of the alveolar wall and hyaline membrane formation. Each item was graded according to the following five-point scale: 0=minimal damage; 1=mild damage; 2=moderate damage; 3=severe damage; and 4=maximal damage. The degree of VILI was assessed by the sum of the scores of items 0 to 16 in five randomly selected high-power fields (HPF, × 400). The average of the total field score was compared among groups.
BALF analysis and NF-κB activity in lung tissue homogenates
For the BAL subgroup, the thorax was opened following exsanguination, and three BAL procedures were performed, each using 1 mL of phosphate-buffered saline (PBS). The retrieval fluid was centrifuged (2,000 g at 4℃) for 10 min and the supernatants were divided into aliquots and stored at -70℃ until measurement of the concentrations of inflammatory cytokines. The concentrations of TNF-α, IL-1β, and IL-6 in BALF were measured by enzyme-linked immunosorbent assay (ELISA) (R&D Systems, Minneapolis, MN, USA). The mean minimum detectable dose of ELISA kits of TNF-α, IL-1β, and IL-6 were 1.88 pg/mL, 2.31 pg/mL, and 1.6 pg/mL, respectively. Nuclear protein from the tissue homogenate subgroup was prepared using a Nuclear Extract Kit (Active Motif, Carlsbad, CA, USA). Activation of the NF-κB p65 subunit was measured in 5 µg of nuclear extracts using an NF-κB p65 ELISA-based transcription factor assay kit (Trans-AM™ NF-κB p65 Transcriptional Factor Assay Kit; Active Motif) (20).
TG2 activity assay
For the TG2 activity assay, lung tissue was homogenized in PBS (pH 7.4) containing a protease inhibitor cocktail (Calbiochem, Darmstadt, Germany), then centrifuged at 14,000 rpm for 15 min at 4℃. The supernatant was stored at -80℃ until analysis. TG2 activity was measured using a chip activity assay (21). Briefly, protein arrays were fabricated by immobilizing fibrinogen on the 3-aminopropyl-trimethoxy-silane surface of well-type arrays. Transamidating activity was determined by probing biotinylated fibrinogen with Cy3-conjugated streptavidin. Amine arrays were prepared by cleaning glass slides (75 × 25 mm) with H2O2/NH2OH/H2O (1:1:5 v/v/v) at 70℃ for 10 min and incubating the slides in 1.5% (v/v) 3-aminopropyltrimethoxysilane solution in 95% ethanol for 2 hr. Teflon tape (75 × 25 mm) with 200 holes (25 × 8) 1.5 mm in diameter was attached to the amine-modified slides. Fibrinogen arrays were fabricated by incubating these well-type amine arrays with 1 µL/well of 50 µg/mL fibrinogen solution in 9.3 mM phosphate buffer (pH 7.4) for 1 hr at 37℃. Fibrinogen arrays were blocked with 3% bovine serum albumin containing 0.1% Tween 20 in phosphate buffered saline (PBS; 8.1 mM Na2HPO4, 1.2 mM KH2PO4, pH 7.4, 2.7 mM KCl, and 138 mM NaCl) for 30 min at 37℃ and washed with 0.1% Tween 20 in PBS and Milli-Q water.
To measure TG2 specific activity, samples and standard mixtures were prepared in 30 µL of buffer containing 40 mM Bis-Tris-HCl pH 6.0, 2 mM CaCl2, 10 mM DTT, 5 mM BAPA, 0.01% Triton X-100, and 140 mM NaCl. For the activity assay, 0.9 µL from each set of reaction mixtures was applied to the fibrinogen arrays described above and incubated at 37℃ for 15 min. Transamidating activity-catalyzed incorporation of BAPA into fibrinogen was probed by incubating the arrays with 10 µg/mL Cy3-conjugated streptavidin at 37℃ for 30 min. Following washing, the arrays were scanned with a fluorescence scanner using a 543-nm laser (ScanArray Express GM; Perkin-Elmer, Waltham, MA, USA). The fluorescence intensities of array spots were measured using a ScanArray Express program (Perkin-Elmer). TG2 specific activity was calculated according to a standard curve obtained from the fluorescence intensities of a TG2-specific standard. TG2 activity for each standard control and experimental sample were measured three times and a mean TG2 activity value was calculated. TG2 activity was expressed as miliunits (mU) per mg of protein (nanomoles per minute per mg of protein) (21).
TG2 quantitative RT-PCR
For RT-PCR, isolated lung tissue was immediately preserved in RNA stabilization reagent (RNAlater, Qiagen, Hilden, Germany) and stored at -20℃ until RNA extraction. Total RNA was extracted from the lung tissue using the Qiagen RNeasy Mini kit (Qiagen) according to the manufacturer's protocol. Total RNA was quantified from the optical density at 260 nm as measured using a NanoDropTM 1,000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). cDNA was synthesized from 1 µg of total RNA in 20 µL using a Transcriptor First Strand cDNA synthesis kit (Roche Diagnostics, Indianapolis, IN, USA) as recommended by the manufacturer. RT-PCR was performed with a LightCycler 2.0 instrument (Roche Applied Science, Mannheim, Germany) using LightCycler FastStart DNA MasterPLUS SYBR Green 1 (Roche Diagnostics). The thermocycler parameters were as follows: 95℃ for 10 min, followed by 95℃ for 15 sec, 60℃ (TG2) and 63℃ (GAPDH) for 1 min, and 72℃ for 40 sec. Real-time PCR signal was monitored by measuring the fluorescence signal during each cycle. The relative expression of each mRNA level was normalized against the corresponding GAPDH level and expressed as a relative change from the control group. Primers for TG2 and GAPDH were synthesized by Bionner Inc. (Daejeon, Korea). The following primer sequences were used in this study: for TG2, forward 5'-TTCGTGTTTGCCGAGGTCAACG-3' reverse 5'-CTTTCTCTGCCAGTTTGTTCAGGTG-3' and for GAPDH, forward 5'-TGTGATGGGTGTGAACCACGAGAA-3' reverse 5'-GAGCCCTTCCACAATGCCAAAGTT-3'.
Administration of TG2 inhibitor
Mice in the Cyst+VILI and Cyst+LPV group received intraperitoneal treatment with 200 mg/kg of cystamine dihydrochloride (Sigma, St. Louis, MO, USA) dissolved in PBS. This dose was found to be effective in LPS-induced acute lung inflammation (15). To determine the optimal pretreatment time, cystamine was administered intraperitoneally to six mice at 36, 24, 12, 6, 4, 2, and 0 hr before LPS instillation and 4 hr of MV. The lowest TG2 activity and TG2 RT-PCR were observed at the 6-hr cystamine pretreatment time (Fig. 1A). Therefore, the Cyst+VILI and Cyst+LPV group mice were pretreated with cystamine at 6 hr prior to LPS instillation and MV and the mice in other groups were pretreated with 200 µL of PBS at 6 hr prior to tracheostomy, LPS instillation, or LPS instillation and MV.
Statistical analysis
All data are expressed as mean ± standard error of the mean (SEM). Statistical analysis was performed using IBM SPSS Statistics for Windows® (Release 20.0; SPSS Inc., Chicago, IL, USA). Inter-group differences were determined by non-parametric Mann-Whitney U and Kruskal-Wallis tests. Statistical significance was defined as P < 0.05.
RESULTS
TG2 activity and TG2 quantitative RT-PCR
TG2 activity (Fig. 1B) was significantly different between groups (P = 0.002 by Kruskal-Wallis test). The LPS (28 ± 1.13 mU/mg of protein) and LPV (29.7 ± 0.84 mU/mg of protein) groups showed higher TG2 activity than the control group (17.9 ± 1.14 mU/mg of protein) (P < 0.05). TG2 activity was significantly increased in the VILI group (41.6 ± 2.16 mU/mg of protein) compared to the LPS and LPV groups (P < 0.05). Cystamine pretreatment decreased TG2 activity significantly in the Cyst+VILI group (32.2 ± 0.92 mU/mg of protein) compared with the VILI group (P = 0.029). However, TG2 activity in the Cyst+VILI group was not significantly different from the LPS and LPV groups (P > 0.05). In the Cyst+LPV group (33.6 ± 1.85 mU/mg of protein), cystamine pretreatment had no additional effect on TG2 activity when compared to the LPV group (P = 0.114).
In the analysis of quantitative RT-PCR of TG2 (Fig. 1C), TG2 gene expression in the VILI group (2.52 ± 0.16 relative change over the control group) was significantly higher than the LPS (1.75 ± 0.07) and LPV (1.63 ± 0.15) groups (P < 0.05). In comparing the VILI and Cyst+VILI groups, TG2 gene expression in the Cyst+VILI group (1.34 ± 0.18) was significantly lower than in the VILI group (P = 0.016). There was no difference in TG2 gene expression in the Cyst+VILI group compared to the LPS and LPV groups (P > 0.05). TG2 gene expression in the Cyst+LPV group (1.21 ± 0.15) was not significantly lower than in the LPV group (P = 0.19).
Inflammatory cytokines and NF-κB activity
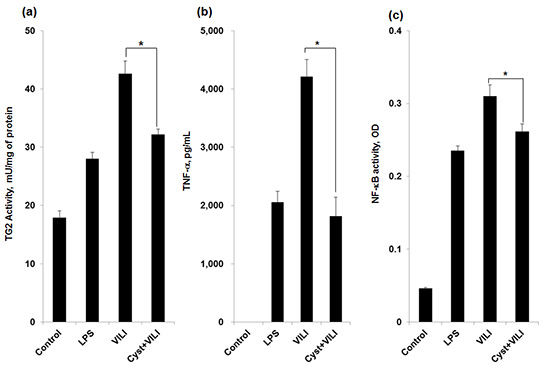
TNF-α, IL-1β, and IL-6 were not detected in the BALF of the control group. The concentration of TNF-α was higher in the VILI group (4,207.17 ± 300.73 pg/mL) than in the LPS (2,053.12 ± 189.26 pg/mL) and LPV (2,348.79 ± 744.5 pg/mL) groups (P < 0.05) (Fig. 2A). The concentration of TNF-α was significantly lower in the Cyst+VILI group (1,814.43 ± 325.14 pg/mL) than in the VILI group (P = 0.016), but was not different from the LPS, LPV, and Cyst+LPV (2,427.79 ± 244.42 pg/mL) groups (P > 0.05). The concentration of IL-1β (Fig. 2B) was higher in the VILI group (432.29 ± 87.99 pg/mL) than in the other groups (P < 0.05). The concentration of IL-1β was significantly lower in the Cyst+VILI group (225.78 ± 47.02 pg/mL) than in the VILI group (P = 0.026), and higher in than the LPV (35.48 ± 7.28 pg/mL) and Cyst+LPV (22.95 ± 6.73 pg/mL) groups (P < 0.05). The Cyst+LPV group had a lower concentration of IL-1β than the LPV group, but this difference was not significant (P = 0.19). The concentration of IL-6 (Fig. 2C) was also significantly higher in the VILI group (2,365.77 ± 275.97 pg/mL) than in the other groups (P < 0.05). Cystamine pretreatment lowered the concentration of IL-6 in the Cyst+VILI group (1,489.11 ± 186.86 pg/mL) more than in the VILI group (P = 0.015). The Cyst+LPV group (611.03 ± 91.01 pg/mL) showed insignificant lower IL-6 concentration than the LPV group (677.71 ± 105.56 pg/mL) (P = 0.429).
NF-κB activity was increased in the VILI group (0.3101 ± 0.0152 optical density, OD) compared with the LPS (0.2349 ± 0.0065 OD) and LPV (0.2465 ± 0.0055 OD) groups (P < 0.05) (Fig. 2D). Cystamine pretreatment significantly decreased NF-κB activity in the Cyst+VILI group (0.2615 ± 0.0107 OD) compared to the VILI group (P = 0.029). NF-κB activity was lower in the Cyst+LPV group (0.2178 ± 0.011 OD) than the LPV group, but this difference was not statistically significant (P = 0.114).
Histopathology and acute lung injury (ALI) score
Histopathologic examination revealed increases in the ALI parameters in the LPS group (Fig. 3B), including increased alveolar capillary congestion, aggregation of inflammatory cells, and thickening of the alveolar wall compared to the control group (Fig. 3A). A higher level of ALI was seen in the VILI group (Fig. 3C) compared to the LPS and LPV groups (Fig. 3E). In the Cyst+VILI group (Fig. 3D), the ALI parameters were attenuated with cystamine pretreatment. In a quantitative comparison of ALI scores (Fig. 3G), the VILI group (12.88 ± 0.29) showed significantly higher scores than the control (2.13 ± 0.29), LPS (8.88 ± 0.58), LPV (8.13 ± 1.29), or Cyst+LPV (8.5 ± 1.09) groups (P < 0.05). Although the ALI score of the Cyst+VILI group (10.88 ± 0.97) was lower than the VILI group, this was not statistically significant (P = 0.105). In comparing the Cyst+VILI group to the LPS, LPV, and Cyst+LPV groups, the Cyst+VILI group had a higher ALI score, but this difference was not statistically significant (P > 0.05).
DISCUSSION
In the present study, TG2 activity and gene expression were significantly increased in a VILI mouse model. The increase in TG2 activity and gene expression was accompanied by increases in the concentration of inflammatory cytokines, NF-κB activity, and ALI score. The pretreatment with cystamine significantly decreased TG2 activity and gene expression, the concentration of inflammatory cytokines, and NF-κB activity, but did not significantly decrease ALI scores.
In terms of managing MV in ALI and ARDS patients, LPV is not always possible due to the heterogeneity of lung injury in some patients (3). A more thorough understanding of the mechanisms that mediate lung injury might permit the development of potential strategies for preventing VILI. TG2 is the most ubiquitously expressed and most studied member of the TG family. This enzyme catalyzes thiol- and calcium-dependent transamidation reactions. TG2 has many functions related to the protection and prevention of injury, as well as tissue remodeling and repair (8). Aberrantly activated TG2 has been implicated in neurodegenerative diseases, atherosclerosis, inflammatory diseases, autoimmune diseases, and fibrosis (9). TG2 is induced by various stressors including LPS, reactive oxygen species, UV, calcium ionophores, retinoic acid, and viral infection (10). Several studies have found that TG2 is involved in the initial phase of inflammation (22, 23). Cytokines and growth factors secreted during the early phases of cell injury regulate TG2 expression. The expression of TG2 is regulated by transforming growth factor-β1 (TGF-β1), TNF-α, IL-1β, and IL-6 (24, 25, 26, 27). Increased TG2 activity induces and exacerbates inflammatory processes via NF-κB activation (10, 11). In studies of ALI models in mice, TG activity was increased, along with TNF-α and IL-6, by the intraperitoneal injection of LPS (28). Intratracheal instillation of LPS also increased TG2 activity along with TGF-β1, TNF-α, IL-1β, IL-6, myeloperoxidase activity, and NF-κB activity (15). However, to date, there have not been any studies examining the role of TG2 in the pathogenesis of VILI, in which injurious MV strategies with high tidal volumes and low PEEP aggravate preexisting acute lung inflammation and induce additional lung injury. Similar to previous studies, this study found that intratracheal LPS instillation resulted in increased TG2 activity and gene expression and increased TNF-α, IL-1β, IL-6, and ALI scores when compared to the control group (15, 28). TG2 activity and gene expression, concentrations of inflammatory cytokines, and ALI scores were significantly elevated in the VILI group compared to the LPS group in response to MV with a high tidal volume and zero PEEP. These results suggest that injurious mechanical ventilation strategies induce aberrant TG2 activation and eventually lead to acute lung inflammation.
TG2-mediated NF-κB activation has been reported to have an important role in inflammatory disease processes. Increases in TG2 activity induce or exacerbate inflammation via NF-κB activation without I-κBα kinase signaling (11). An increase in TG2 activity reduces free I-κBα in the cytosol via I-κBα polymerization, resulting in the translocation of free NF-κB into the nucleus. In turn, TG2 can be directly induced by NF-κB activation as the TG2 promoter contains a NF-κB binding motif (29). In this study, the LPS-induced inflammatory reaction and the increase in TG2 activity and gene expression were also accompanied by an increase in NF-κB activity. Injurious MV strategies also exacerbated inflammation and increased TG2 activity and gene expression and concentrations of NF-κB. However, this study was limited by the fact that I-κBα polymerization was not examined.
The therapeutic possibilities of TG inhibitors have been evaluated in various disease models. TG inhibitors have the potential to reverse inflammatory damage in brain injury, conjunctivitis, and uveitis (10). In the present study, pretreatment with cystamine, which is a unique inhibitor of TG2 with multiple inhibition mechanisms (30), significantly attenuated TG2 activity and gene expression in mice which had experienced injurious MV. Furthermore, cystamine decreased concentrations of inflammatory cytokine and the activity of NF-κB. Cystamine pretreatment resulted in a trend towards decreased ALI scores; however, the changes were not statistically significant, suggesting that TG2 inhibition was not sufficient and has limitations in the histopathologic reduction of VILI. The LPV strategies showed a protective effect by decreasing TG2 activity and gene expression, concentrations of inflammatory cytokines, NF-κB activity, and ALI score. The addition of cystamine pretreatment to LPV did not significantly change the parameters compared with the LPV group. TG2 gene expression, IL-1β and IL-6 concentrations, and NF-κB activity showed a downward trend in the Cyst+LPV group, more than in the LPV group. There may be limited benefits from the addition of TG2 inhibition to LPV. Cystamine pretreatment only with LPS instillation (Cyst+LPS) group was not experimented in the present study. According to the study of the time response of TG2 activity after 0.5 mg/kg of LPS intratracheal instillation (15), TG2 activity was reported to be highest 36 hr post LPS instillation. The increase of TG2 activity 3 and 6 hr post LPS instillation was modest compared to the control group. It was judged to be inappropriate to examine TG2 activity and the effects of TG2 inhibition at the time of 4 hr after LPS instillation without additional VILI. In the preliminary study, the TG2 activity was experimented in the model using an injurious MV strategy alone without LPS instillation. However, increase in the TG2 activity was not significant in this model. This study was therefore performed in the model with both LPS instillation and injurious MV strategy, which might be more clinically relevant with ARDS.
There are several limitations in this study. The most important limitation is omission of data about pulmonary dynamics and physiologies. Under the mechanical ventilator setting with high tidal volume, intrinsic PEEP could develop and influence pulmonary dynamics of the animal. However, pulmonary dynamics could not be measured in this experiment, and other data such as arterial partial pressure of oxygen and lung water content which reflect the degree of lung injury were not measured. In the inflammatory disease, the pro/anti-inflammatory balance is important. Therefore, measurement of anti-inflammatory cytokines could be necessary in observing the balance. However, none of anti-inflammatory cytokines was examined in this study.
The aberrant activation of TG2 plays an important role in the inflammatory processes that underlie the development of VILI. The inhibition of TG2 had therapeutic effects on the inflammatory parameters of VILI in mice, suggesting potential clinical usefulness in the prevention and treatment of VILI in ARDS patients.


 XML Download
XML Download