

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Alkali therapy in chronic renal failure (CRF) ameliorates the progression of kidney injury (1, 2, 3, 4) and better preserves renal function (5). Several mechanisms behind its effects have been evaluated, including the reduction of inflammation (6, 7), cytokines (8) and endothelin production (5). In our previous study, bicarbonate therapy downregulated apical Na/H exchanger type 3 (NHE3) expression in CRF (9). Luminal NHE3 is a significant transporter for renal Na+ and HCO3- absorption (10) and could be a main therapeutic target of alkali therapy for renal disease progression (9).
Pressure-natriuresis is a classical concept that describes the association between blood pressure and sodium balance; kidney damage alters pressure-natriuresis and the positive sodium balance. Salt sensitivity, which shifts rightward on the pressure-natriuresis curve, is a risk factor for cardiovascular outcomes in hypertensive patients (11). Salt sensitivity may increase not only intraglomerular pressure but also an additional risk of glomerular damage and microalbuminuria (12, 13, 14); therefore, preserving the pressure-natriuresis relationship may be a preventive method for the progression of kidney injury. In addition, metabolic acidosis is observed in salt-sensitive hypertension (15), and NHE activity in platelet is increased in some essential hypertensive patients (16). In animal studies, altered NHE3 expression has been associated with the change in salt sensitivity (17). However, few reports have investigated the effects of alkali therapy on salt sensitivity and kidney injury in CRF. We used a remnant kidney model to evaluate the beneficial effects of dietary sodium citrate on salt sensitivity and on ameliorating renal disease progression in rats.
MATERIALS AND METHODS
Twenty-four 6-week-old male, specific pathogen-free Sprague-Dawley rats, purchased from Orient Bio Inc. (Seongnam, Korea) were given ad libitum access to water. Experimental CRF was induced by the sequential excision of approximately two-thirds of the left kidney and total right kidney (9, 18). After 5/6 nephrectomy, the CRF animals were randomly distributed to the standard diet group (Agribrands Purima Korea, Seongnam City, Korea), sodium chloride (NACL)-treated group or sodium citrate (NACT)-treated group. The NACL-treated CRF rats were fed a 20% casein diet with 174 µM/g NACL (DYET #113811, Dyets Inc., Bethlehem, PA, USA), and the NACT-treated CRF rats were fed a 20% casein diet with 174 µM/g NACT (DYET #103316). Sham-operated rats were divided into 3 groups and administered the same diet. All operated animals were sacrificed postoperatively at week 4.
We measured blood pressure using the tail-cuff plethysmography method (IITC Life Sciences, Woodland Hills, CA, USA). We collected 24-hr urine samples in the metabolic cages on the day before sacrifice, and blood samples from the abdominal aorta at the time of sacrifice for measuring urea nitrogen, creatinine, osmolality, and electrolytes. We performed biochemical tests using an automatic chemistry analyzer (Hitachi 7070; Hitachi, Tokyo, Japan). We used indirect ion-selective electrode methods for electrolyte levels, and the enzyme method for total CO2 levels. We measured plasma and urine osmolality with a cryoscopicosmometer (Osmomat 030-D-M; Gonotec, Berlin, Germany). We calculated the glomerular filtration rate (GFR) using the average of the urea clearance and creatinine clearance (10).
Total extracted protein concentration was measured using the bicinchoninic acid protein assay method (BCA Reagent Kit; Sigma, St. Louis, MO, USA). Briefly, samples were fractionated by SDS-PAGE. After transfer onto the Protran Nitrocellulose Transfer Membrane (Whatman, Gassel, Germany), the blots were probed with the primary antibodies (9, 10). Previously characterized polyclonal antibodies were used for semiquantitative immunoblotting and immunohistochemistry, as described in the previous studies (9, 10). In addition, affinity-purified polyclonal antibodies against sodium dicarboxylate transporter (NaDC1, sc-23539, Santa Cruz Biotechnology, Inc., Santa Cruz, CA) were used.
As previously described (9, 10), the glomerulosclerosis index and the tubulointerstitial damage index were evaluated using a semiquantitative scoring system, which were performed in a blinded fashion. The immunostaining procedure was performed according to the protocol of the Dako Cytomationkit (Envision + Dual Link System-HRP; Carpinteria, CA, USA) (9, 10). Multiple fields of each kidney sections were photographed with 40× objective lens using a light microscope (Axioskop40, Carl Zeiss, Goettingen, Germany), and staining intensity was quantified using region in Axiovision software (v4.8.2).
Tissue endothelin (ET)-1 levels were measured by quantitative enzyme immunoassay using QuantiGlo ET-1 ELISA (QET00B, R&D Systems, Minneapolis, MN, USA).
RESULTS
Physiologic data, salt sensitivity, and pathologic data
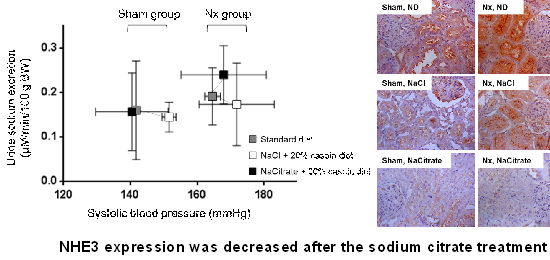
As shown in the Table 1, the NACT-treated CRF group had greater levels of the serum total CO2 than the NACL-treated group 4 weeks after surgical reduction of the kidney mass. Regarding the pressure-natriuresis relationship (Fig. 1), the point of pressure-natriuresis in the SD-treated animals was shifted to the right after nephrectomy. Also, the point was additionally shifted to the right in the NACL-treated rats, compared to the SD-treated groups, regardless of whether they were sham-operated or nephrectomized. In the CRF groups, the systolic blood pressure in the NACT-treated group showed a tiny reduction, compared with the NACL-treated group. The NACT diet triggered an increased urinary sodium excretion rate, with elevated blood pressure. Conversely, the urinary sodium excretion rate in the NACL-treated CRF group was not increased, although systolic blood pressure was increasing; the pressure-natriuresis relationship was shifted to the right.
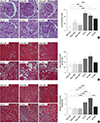
At week 4, a small rise in the GFR in the NACT-treated CRF group was found compared to the NACL-treated CRF group, though the GFRs of both CRF groups were lower than those of SD-treated CRF group (Table 1). The glomerulosclerosis index and cortical tubulointerstitial damage index in the NACT-treated CRF group were relatively modest compared to the NACL-administrated CRF group, although not statistically significant (Fig. 2). The medullary tubulointerstitial damage index in the NACL-administrated and SD-fed CRF group was twice that of the sham-operated group; however, the NACT treatment abrogated the kidney injury caused by nephrectomy (Fig. 2).
Change of renal transporters
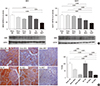
After nephrectomy, NHE3 expression in the NACT-treated group had fallen compared to the NACL-treated group. Similarly, immunohistochemistry indicated weaker expression for NHE3 in the NACT-treated group than the other groups (Fig. 3).
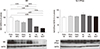
The densities of NKCC2, NCC, ENaCs and H-ATPase were decreased after nephrectomy compared to the sham-operated groups; however, no differences between NACL- and NACT-treated CRF groups were shown. SGLT1 and pendrin expressions did not differ in the six groups (data not shown). NBC expression decreased after the NACT treatment in both the CRF and sham-operated groups; however, no difference between the NACL- and NACT-treated groups was found (Fig. 4). The Na-K-ATPase expression did not differ in all of the six groups (Fig. 4).
The renal ET-1 levels increased after nephrectomy with the SD and NACL treatments. ET-1 in the NACT-treated group was slightly lower than that in the NACL-treated group although not statistically significant (Fig. 5).
DISCUSSION
The study data demonstrated that dietary NACT in nephrectomized models may improve the pressure-natriuresis relationship and ameliorate kidney damage, compared to the NACL diet. The underlying mechanisms of these beneficial effects can be associated with a preserved pressure-natriuresis relationship, which may be caused by the decreased expression of NHE3, with the correction of metabolic acidosis.
Some studies have indicated that salt sensitivity is associated with substantial kidney damage (12, 13, 14). Additionally, salt sensitivity is a major risk factor for kidney injury (12, 13, 14) and cardiovascular events (11). In the current study, alkali therapy contributed to the reverse of the pressure-natriureis points, which shifted to the right after the NACL treatment. In alkali-treated CRF rats, NHE3 expression was dramatically suppressed; this suppression may lead to increased sodium excretion. It may help to maintain pressure-natriuresis and decreased blood pressure despite the renal mass reduction. Other alkali therapies, such as sodium bicarbonate, also reduced NHE3 expression in CRF models (9); these beneficial effects of alkali therapy in CRF may improve pressure-natriuresis relationship.
Sharma et al. showed that NACL increases blood pressure in salt-sensitive subjects whereas NACT does not (15). Additionally, the authors demonstrated that the elevated blood pressure induced by NACL is associated with metabolic acidosis and that NACT can correct acidosis in salt-sensitive subjects (15). This result is fully consistent with our data. We demonstrated altered NHE3 expression and the shift of pressure-natriuresis. The proximal tubule reabsorbs two-thirds of the filtered sodium. As blood pressure changes, this fraction of reabsorption is appropriately adapted. Zhang et al. showed that more than half of the apical NHE3 immunoreactivity was redistributed to membranes after blood pressure increased and that all NHE3 returned to the original distribution levels after blood pressure normalized (17); therefore, NHE3 may affect the regulation of sodium reabsorption in response to fluctuations in sodium or blood pressure (19).
Acid retention during kidney failure induces ET production, which leads to progressive GFR decline (20). ET-1 plays a role in pathophysiology in the development and maintenance of salt sensitivity, and an ET receptor antagonist may be useful in treating salt-sensitive hypertension (21). Previously, apical NHE3 expression had fallen in the bicarbonate-treated group with a slight reduction of ET levels (9). In the current study, NACT therapy reduced the kidney levels of ET in CRF rats, but it was not statistically significant, which suggests that ET-1 may not be a main mechanism, although reducted ET might influence the relationship between pressure-natriuresis and alkali therapy. In addition, alkalization can directly influence on the downregulation of apical NHE3 activity in the renal proximal tubule unlike other sodium transporters, such as SGLT1, NKCC2, NCC, and ENaC (22). Interestingly, Na-K-ATPase was not decreased, contrary to the previous study, which described decreased Na-K-ATPase after bicarbonate therapy (9, 23). This result may suggest that citrate may not affect NHE3 via Na-K-ATPase. Unlike NHE3 expression, H-ATPase or NBC abundances, which are crucial mediators of acid-base balance, may not be connected with citrate treatment in CRF; therefore, NHE3 will be one of the major targets of alkali treatment based on citrate.
Citrate is reabsorbed in the renal proximal tubule by a sodium-dicarboxylate cotransporter (NaDC) and produces bicarbonate. Aruga et al. indicated that metabolic acidosis increased NaDC abundance and that alkali loading did not alter NaDC expression (24). In addition, citrate can be metabolized or shuttled into the mitochondria where it enters the citric acid cycle (25). It may be possible that the beneficial effect of citrate, such as improvement of mitochondrial dysfunction in chronic renal failure, may assist in ameliorating kidney injury, a mechanism that differs from alkali therapy based on bicarbonate.
In conclusion, we found that citrate therapy in CRF may have positive effects on salt sensitivity and ameliorate the progression of kidney injury, and that these effects could be related to the altered expression of NHE3 and correction of metabolic acidosis. These results suggest that alkali therapy might be applied to other salt-sensitive pathologic conditions as well as CRF.



 XML Download
XML Download