

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Herpes simplex virus type 1 (HSV-1) is a member of the herpesvirsus family, which infection commonly involves recurrent mucocutaneous manifestations (1). HSV-1 could also cause illness in a much severe form; ocular herpes could cause adult blindness, furthermore induce sporadic encephalitis, usually resulting in severe morbidity or mortality (2). Moreover, HSV-1 is one of the leading infectious viral pathogens found in immunocompromised hosts, such as transplant recipients. HSV-1-induced tissue damage comprises of mononuclear cell infiltration, perivascular inflammation, as well as various cytopathogenic changes including intracellular inclusion bodies and syncytial formation (3). Especially, syncytial formation is a characteristic pathologic response seen in herpesvirus or human immunodeficiency virus type 1 (HIV-1) infections (3, 4).
This fusogenic activity in HSV-1-infected cells requires the presence of specific glycoproteins in the virion envelope, including glycoprotein B (gB), gD, gH, and gL (5-8). Syncytial formation starts with the binding of virus-infected cells to uninfected cells through viral glycoprotein-receptor interaction, followed by cell-to-cell fusion to form a multinucleated giant cell, the syncytium. This eventually ruptures to cause cell death. It is noteworthy that patients infected with syncytium-inducing (SI) HIV-1 variants have a more rapid deterioration in their course than those infected with non-SI variants (9, 10).
In immunocompromised hosts infected with HSV-1, concurrent infections by other microorganisms, including other viruses are not uncommon. Human cytomegalovirus (HCMV) is another important causal agent in opportunistic infection, which can induce comparable cytopathogenic effects in infected cells (11). There have been numerous reports of co-infection of HSV-1 and HCMV in immunocompromised patients (12-14). The clinical outcome of herpesvirus co-infection is most appreciated in follow-up studies after transplantation. For example, grafts infected with both HSV-1 and HCMV results in shorter graft and patient survival than grafts with either single viral infection (14). However, there are no study yet which investigates the pathology in HSV-1, HCMV co-infected tissue or organs.
We hypothesized that in the setting of active HSV-1 and HCMV co-infection, HCMV could exacerbate HSV-1-induced cytopathogenic changes in HSV-1 permissive cells. In particular, we focused on the HCMV gene products from the major immediate early (IE) gene. By utilizing a U373MG cell line expressing HCMV IE1 protein (15), we investigated the function of HCMV IE1 protein in modulating HSV-1 induced syncytial formation.
MATERIALS AND METHODS
Cell lines and virus
We utilized cell line UMG1-2 that constitutively expresses HCMV IE1 protein in U373MG cells (ATCC HTB 17) (15). U373MG cells transfected with the empty retrovirus vector LNCX2 vector (Clontech, Palo Alto, CA, U.S.A.) was used as the control. Cells were cultured in Dulbecco's modified Eagles medium (GIBCO, Grand Island, NY, U.S.A.) with 10% fetal bovine serum (GIBCO), 100 U/mL penicillin and 100 U/mL streptomycin (Life Technologies, Carlsbad, CA, U.S.A.) in a 37℃ incubator with 5% CO2. HSV-1 MacIntyre (ATCC VR-539) was used in the experiment.
Reagents
Neutralizing anti-HSV-1 monoclonal antibody, MHSVI116 (2 µg/mL) was used as previously described (16). Other reagents were used in the following concentration: ganciclovir (40 µM), mitomycin C (1 µg/mL), nocodazole (10 µg/mL), N-tosyl-1-phenylalanine-chloromethyl ketone (TPCK) (10, 100 µM), emodin (1, 10 µM) and roscovitine (2-100 µM). All reagents were purchased from Sigma (St. Louis, MO, U.S.A.). Concentrations of the drugs were determined by appraisal of former reports of drug treatment in U373MG cells or in studies utilizing HSV-1 (15, 17-19). Drugs were prepared in 100× concentrations.
HSV-1 infection and syncytial formation
Ten thousand cells were plated on each compartment of an 8-chamber slide (Sigma) and cultured for 24 hr. HSV-1 stock (2.0×106 plaque forming unit/mL) was diluted 10 fold from 10-1 to 10-4 in volume. UMG1-2 and its control in each compartment were infected with 100 µL of HSV-1 for 1 hr followed by washing phosphate-buffered saline (PBS) and addition of 100 µL of media. Slides were fixed with 4% paraformaldehyde-PBS after 6, 12, 24, 48, and 72 hr and stained with hematoxylin & eosin. The size of each syncytium was evaluated by visible nuclei numbers per syncytium under a light microscope (8). Syncytium with 5 or more nuclei was enumerated in five low-power fields. Experiments were repeated three times.
Immunohistochemistry
Cells were infected with 0.2 multiplicity of infection (m.o.i.) of HSV-1 for 1 hr and were added with media after washing with PBS. Immunohistochemistry was done with the following protocol; cells were fixed with 4% paraformaldehyde-PBS for 1 hr and then permeabilized with 0.5% Nonidet P-40 (Sigma)-PBS for 5 min. After washing, wells were blocked with 2% skim milk for 30 min, and then treated with MHSVI116 antibody for 1 hr at room temperature. Peroxidase-conjugated anti-mouse IgG (Jackson Immunoresearch, West Grove, PA, U.S.A.) in 2% skim milk was added for 1 hr at room temperature. Finally, wells were treated with 0.2% diaminobenzidine (Sigma) and 0.005% hydrogen peroxide in PBS for 20 min. Reaction was stopped with distilled water and the plates were observed under an inverted microscope.
Drug treatment
Cells were concomitantly treated with the drug of interest with HSV-1 infection (0.2 m.o.i.). Culture media with or without dimethyl sulfoxide (DMSO, Sigma) was used as controls. After infecting cells with HSV-1 for 1 hr, cells were washed with PBS, and then each drug concentration was maintained for 24 hr. Cells were fixed and immunohistochemistry was performed as described, followed by enumeration of syncytia (nuclei ≥5) under an inverted microscope. Experiments were repeated three times.
RESULTS
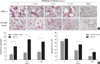
Enhanced syncytial formation in U373MG cells expressing HCMV IE1 protein
Both UMG1-2 and U373MG control cells formed syncytia after HSV-1 infection. Syncytial formation was negligible at 6 hr post-infection even in high HSV-1 concentrations but noticeable at 12 hr at 2 m.o.i. (data not shown). At 24 hr, we could appreciate a clear m.o.i.-dependent syncytial formation by HSV-1 (Fig. 1A). The syncytial formation effect was enhanced in UMG1-2 cells compared with the U373MG control, especially the size of the syncytium. The nuclei number per syncytium was significantly higher in UMG1-2 cells than in U373MG control cells (Fig. 1B). The number of syncytia in the two groups was not different at 2 m.o.i., as all sizes of syncytia were included. With ongoing fusogenic activity in both groups of cells, syncytia enumeration was difficult in longer periods of infection (48, 72 hr) due to cell lysis, especially in UMG1-2 cells in which cell lysis was detected earlier, at a lower m.o.i. due to robust syncytial formation (data not shown).
Drug effects on syncytial formation
For further characterization of the enhanced cytopathogenic changes in UMG1-2 cells, cells were treated with blocking antibody or inhibitors pertinent to viral multiplication and intracellular signaling associated with herpesvirus infection (Fig. 2A). First, MHSVI116, a neutralizing antibody against HSV-1 gB, completely abrogated syncytial formation by HSV-1 in both UMG1-2 and U373MG controls. Nocodazole was used to impair microtubule-mediated transport of HSV-1 capsid to the nucleus (20). It markedly suppressed syncytial formation in both cell lines as expected. Syncytial formation was also compromised in both cell lines by blocking viral DNA synthesis with ganciclovir or mitomycin C. After treatment of cyclin-dependent kinase (cdk) inhibitor roscovitine, both UMG1-2 and U373MG controls showed decreased syncytial formation (Fig. 2B). Importantly, the enhanced cytopathogenic response in UMG1-2 was diminished by roscovitine. Moreover, nuclei numbers per syncytium were significantly lower in both cell lines treated with roscovitine (media vs. roscovitine-treated UMG1-2 cells ([mean±SD] 27.4±5.8 vs. 12.1±3.5, p=0.001). Other inhibitors such as tyrosine kinase inhibitor, emodin, and NF-κB inhibitor, TPCK, did not suppress syncytial formation in either cell lines after infection.
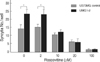
Dose-dependent inhibition of syncytial formation by roscovitine
To further investigate the dose response of roscovitine in HSV-1-infected cells, we next compared syncytial formation in different concentrations of roscovitine (Fig. 3). Roscovitine inhibited syncytial formation in both HSV-1-infected cells in a dose-dependent manner at 24 hr post-infection. With increase concentration of roscovitine, enhanced syncytial formation in HSV-1-infected UMG1-2 cells was effectively neutralized.
DISCUSSION
HSV-1 infection with co-infection with other viruses, could cause severe illness in immunocompromised hosts (14). By utilizing a HSV-1 permissive U373MG cell line constitutively expressing HCMV IE1 protein, we demonstrated enhanced syncytial formation in IE1 protein expressing UMG1-2 cells after HSV-1 infection. Furthermore, we showed that along with potent viral replication inhibitors and neutralizing antibodies, roscovitine could effectively inhibit the augmented the cytopathogenic effect. To better control the variability produced by the complex biology of co-infection models, we postulated that HCMV major IE gene products could be a major factor modulating HSV-1 pathogenecity in host cells co-infected with both herpesviruses. This hypothesis was supported by in vitro studies demonstrating HSV-2 reactivation by HCMV in the very early stages of HCMV super-infection (21, 22).
HCMV IE1 protein, one of the two (IE1 and IE2) transcription factors of the major IE promoter, is the most abundantly expressed viral protein in the immediate early phase of HCMV-infected permissive human cell lines (23, 24). HCMV IE1 protein is expressed shortly after HCMV infection and its level increases steadily through 72 hr (25). Moreover, HCMV infection induces dominant expression of IE1 protein compared with IE2 protein in many cancer cell lines such as Saos-2 and U373MG (15). In fact, HCMV IE1 protein helps expression of viral and cellular promoters synergistically with IE2 protein (26). Interactions have been reported between HCMV IE1 protein and other transcription factors, such as SP-1 and CTF-1, or the TFIID complex (27-29). Especially, HCMV IE1-TFIID interaction leads to inhibition of apoptosis induced by tumor necrosis factor in vitro (30, 31).
The functional role of HCMV IE1 protein in host cell growth has been previously described in several transfection studies (32-35). HCMV IE1 protein was first known to interact with retinoblastoma susceptibility gene-related p107 protein to overcome p107-induced cellular growth arrest (33, 34). Subsequent studies revealed that HCMV IE1 but not IE2 protein, could bind to p107 and facilitate cyclinE/cdk2 activation (35). Furthermore, HCMV IE1 protein induces resistance to apoptosis in U373MG cells by cdk2 accumulation (15). Cdks are required for replication of many viruses, including HIV-1, HSV-2, HCMV, and varicella-zoster virus (36-40). Among pharmacological cdk inhibitors, roscovitine has already been investigated as a potential anticancer drug for disrupting the cell cycle (41). The emerging interest in roscovitine is its antiviral effect, such that it specifically inhibits genes in viruses such as HIV-1, HSV-2, HCMV, and varicella-zoster virus (42, 43). In HSV-1-infected cells, roscovitine blocks accumulation of mRNAs encoding specific viral IE and early genes, inhibits viral DNA synthesis, and alters posttranslational modification of HSV-1 IE protein (38, 44-47). In general, roscovitine is likely to inhibit HSV-1-induced syncytial formation by interfering viral replication. Considering the significant reduction in syncytial size, we further speculate roscovitine may also have affected late expression of viral proteins, including viral glycoproteins necessary for syncytial formation.
Inhibitors of NF-κB, and tyrosine kinsase we used did not affect syncytial formation in HCMV IE1 protein expressing cells. Interestingly, previous studies showed that HCMV IE1 protein selectively induce both NF-κB complexes and its activity (48-50). In addition, HSV-1 infection also results in intranuclear translocation of NF-κB (51). However, our results show that NF-κB signaling is not the major pathway that HCMV IE1 protein contributes to the phenotype of enhanced syncytial formation by HSV-1. As for the tyrosine kinase inhibitor, emodin, we were unable to detect any evidence of emodin affecting syncytial formation, in spite of reported antiviral effects (19, 52).
There are yet limitations to indicate HCMV IE1 protein to be the essential molecule in vivo modulating increased cytopathogenecity in HCMV, HSV-1 co-infected cells. Our model is based on the constitutive expression of HCMV IE1 protein in host cells, disregarding the life cycle and other gene products of HCMV. Furthermore, previous in vitro studies demonstrate functional conservation in viral gene products of both HCMV and HSV-1 (53-55). Thus, it is possible that other HCMV gene products could also potentiate cytopathogenicity induced by HSV-1, especially by facilitating viral replication (56). Further investigations will include identifying genes modulated by HCMV IE1 in the syncytium, especially genes in viral glycoprotein synthesis.
In summary, this is the first study to implicate the potential contribution of HCMV IE1 protein to enhance cytopathogenic responses in active HSV-1 infection. In addition to known potent inhibitors of viral replication, roscovitine was able to effectively suppress HCMV IE1 protein-induced cytopathologies, supporting its effective anti-viral activity.

 XML Download
XML Download