

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Since estrogen plays its role through its binding to estrogen receptors (ERs), ER-α and ER-β, the morphological information on the distribution of ERs became imperative in understanding the effect of estrogen on various intracellular molecular dynamics. Among various organs exhibiting ER immunoreactivities (IRs), the central nervous system (CNS) was intensively investigated because estrogen was recently revealed to be crucial for neuroprotective functions (1-3). These novel functions of estrogen in the CNS were reported to be related to the presence of ERs within the cytoplasm (4-6), which were quite different from those of previously recognized ERs in the nuclei of cells (7-10). Among the neural cells showing possible estrogen-related neuroprotective effect, astrocytes were important candidates for the function because ER-α IRs were shown to be intensely localized within astrocytes of affected brain regions in some pathological states. ER-α was exclusively localized within astrocytes exhibiting glial fibrillary acidic protein (GFAP) or vimentin IRs, the markers for reactive astrocytes against various injuries to the brain (11). In this regard, reactive astrocytes could be a direct target for estrogens working through its binding to ERs within the cells. Taken together, ER-α IRs were mainly observed in neurons in normal state but ER-α IRs shifted into astrocytes in some pathological states.
However, we found out that ER-α IRs were expressed differentially even among GFAP-positive astrocytes, suggesting that the expression of ER-α IRs might affect the expression of GFAP and even the process elongation of astrocytes. The findings of this study also suggests that the expression pattern of ER-α IRs in reactive astrocytes under neuropathlogical conditions could be more complicated than have been suggested by previous studies.
CASE REPORT
The subject of the current case, human female neonate, was born at 41 weeks of gestation by cesarean section. After she died on the sixth postnatal day, autopsy was performed. The postmortem examination showed various anomalies of cardiovascular, digestive, and urogenital systems. The anomalies included patent ductus arteriosus, pyrolic stenosis of the stomach, exstrophy of the urinary bladder, urethral atresia, omphalocele-containing urinary bladder, vaginal atresia (complete obstruction), and anorectal agenesis with rectovesical fistulas. However, no specific morphological anomalies were detected in the brain by gross and microscopic examinations. The cerebellar tissue sample was taken and immersed overnight in the freshly prepared 4% paraformaldehyde (PFA)-containing phosphate-buffered saline (PBS). Subsequent tissue processing and immunohistochemical study were performed in accordance with our previous method (12). The primary antibodies used in the present study were rabbit anti-human ER-α antibody (1:1,000, Santa Cruz, CA, U.S.A.), mouse anti-GFAP antibody (1:500, Biogenix, CA, U.S.A.), mouse anti-OX42 antibody (MCA275G, SeroTec, U.K.), and mouse monoclonal anti-165 kDa neurofilament (1:500, Developmental Studies Hybridoma Bank, Iowa, U.S.A.) antibody. The secondary antibodies used for fluorescence microscope were alexa fluor 555, and alexa fluor 488 antibodies against rabbit and mouse (Molecular Probes, Eugene, OR). DAPI (1:5,000, Molecular Probes) was used for nucleus staining.
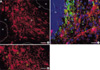
Intense ER-α IRs were observed mainly within the white matter than those in the gray matter including granular and molecular layers (Fig. 1A, B). To rule out the possibility that ER-α IRs were expressed by cerebellar neurons, double immunohistochemical staining was performed with anti-ER-α and anti-165 kDa neurofilament antibodies. Anti-165 kDa neurofilament IRs were not co-localized in the cell processes exhibiting ER-α IRs (Fig. 1C).
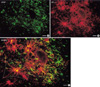
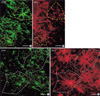
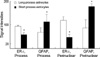
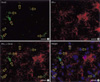
The types of ER-α-positive cells were also identified by a double immunohistochemical study with both anti ER-α and GFAP antibodies. The majority of the cells expressing ER-α IRs showed GFAP IRs (Fig. 2A to C). And there might be some differences in the process length among the reactive astrocytes showing both GFAP and ER-α IRs. The reactive astrocytes with intense ER-α IRs exhibited relatively long processes and relatively weak GFAP IRs (Fig. 3). To confirm this pattern with a statistical method, the reactive astrocytes exhibiting GFAP IRs were classified into two groups; the one with long processes (>20 µm of process length) and the other with short processes (below 20 µm of process length). The intensities of GFAP IRs and ER-α IRs were also differentially measured in perinuclear or process areas within the cells in each group. The signal intensities were measured in 100 different photographed fields using Image program (Scion Corporation, Frederick, MD, U.S.A.). All densitometric data were expressed as mean density, i.e. the average of the gray values of all pixels within the measured area (maximum level is 256). The acquired data were analyzed by Student t-test, and a p value less than 0.01 was deemed statistically significant. The final data were presented as means±S.D. In this quantitative study, we statistically confirmed that GFAP immunoreactive astrocytes with long processes expressed much intense ER-α IRs than those with short processes (Fig. 4). ER-α IR cells were not reactive for a microglia marker, OX-42 (Fig. 5).
DISCUSSION
It was well known that ER-α was mainly expressed by neurons in the brain without any neuropathological findings (13-15). On the other hand, we showed that much intense ER-α IRs were observed mainly within astrocytes of the white matter than neurons in the gray matter of the neonatal cerebellum with multiple congenital anomalies of the present case. Since ER-α was known to be localized within astrocytes in cases of excitotoxic injury or Alzheimer disease (11, 16), the current study could be well matched with previous reports. Therefore, even though the exact neuropathology of the current case was not clearly revealed in this study, the abnormal expression pattern of ER-α, the decreased intensities of neuronal ER-α IRs and increased intensities of ER-α IRs in astrocytes, could suggest that the neonatal brain might be under abnormal neurological conditions, which might finally induce the astrocytes into reactive states.
Though we could not be sure if estrogen directly affect the elongation of the astrocyte processes, it should be also considered that estrogen might inhibit GFAP transcription in astrocytes, which in turn down-regulate the proliferation of reactive astrocytes after injury (17).
As to the another possible cause of process elongation of ER-α-positive reactive astrocytes, the previous report on the effect of estrogen on promoting neurite sprouting under nerve growth factor (NGF) stimulation (18) deserves consideration. According to them, estrogen treatment markedly enhanced NGF-stimulated neurite outgrowth of PC12 cells that were stably transfected with the full-length rat ER-α gene (PCER). Since the estrogen effect on the neurite sprouting was thought to be correlated with the modulated expression of various cytoskeletal proteins (18), we could suggest the possibility that estrogen might also affect the cytoskeleton dynamics through ER-α during the process elongation of reactive astrocytes.
Although we could not clearly elucidate the cause of process elongation in ER-α IR astrocytes currently, the present study demonstrated for the first time that among reactive astrocytes under abnormal conditions, cells that express ER-α intensely had much longer processes, which might have been caused by the decrease in GFAP expression or inducing effect of estrogen on cytoskeleton dynamics. However, since our findings were from a single case under unknown pathological stimuli, our hypothesis should be considered as a tentative suggestion until the general mechanism related to the current changes could be further elucidated in the forthcoming animal experiments.
 XML Download
XML Download