

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Netrin family proteins play essential roles in the development of the axonal pathway and neuronal migration during embryonic development of the vertebrate nervous system (1-3). The commissural neurons in the embryonic spinal cord project their axons to the ventral side of the spinal cord, and cross midline to give rise to commissural axons (4). Netrin is secreted from the floor plate of the spinal cord. They attract the developing commissural axons (4, 5), and the commissural neurons express the netrin receptor, Deleted in Colorectal Cancer (DCC) (6). DCC is the receptor that signals a chemoattractive response to netrin (6, 7). Netrin also repels axons from trochlear motor neurons (8). Thus, netrin is a bifunctional molecule, and some of the repulsive actions of netrin appear to be mediated by other receptors, namely, the Unc5h family of receptors (9, 10). Thus far, four members of the Unc5h family, Unc5h1-4, have been reported (11, 12).
It was recently reported that netrin and its receptors are expressed in the adult nervous system (12-14). For example, the neuronal and non-neuronal cells in the adult spinal cord express both netrin and the receptors, that is, DCC and Unc5h (12, 14). In addition, netrin and the netrin receptors continue to be expressed in the adult optic nerve and retina (15). Although the expression of netrin and its receptors in the adult nervous system indicates that netrin is possibly involved in the function of adult neurons, it remains uncertain whether netrin has any functions in the axonal growth, particularly in the regenerative growth, of mature neurons. Here we observed that the adult dorsal root ganglion (DRG) expresses mRNAs of three members of the Unc5h family and that netrin has an inhibitory effect on the regenerative axonal growth of cultured DRG neurons.
MATERIALS AND METHODS
Primary cultures
For explant cultures, lumbar DRGs were removed from 40-50 days old male Sprague-Dawley rats. The rats were used after our college ethics committee approved the protocol, stating that it fulfilled the animal care guidelines established by the Korean Academy of Medical Sciences. After the dissection of a DRG into 3-4 explants under a stereomicroscope, the explants were cultured in Matrigel (BD Bioscience, San Jose, CA, U.S.A.) in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA, U.S.A.) supplemented with 10% fetal bovine serum (Invitrogen). Recombinant netrin-1 (500 ng/mL, R&D Systems, Minneapolis, MN, USA) was treated for 36-48 hr. After fixation with 4% paraformaldehyde (PFA), the axons were stained with anti-β-tubulin (Tuj1) antibody (1/2,000, Covance, Irvine, CA, U.S.A.) as previously described (16). Alexa 488-conjugated anti-mouse IgG (1/800, Promega, Madison, WI, U.S.A.) was used as the secondary antibody. The fluorescent signals were examined under a laser scanning confocal microscope (Carl Zeiss, Hamburg, Germany), and the signals were analyzed using the KS 400 image system (Carl Zeiss). Quantification of the axon projections from the DRG explants was carried out; the number of explants (n) in each group in an experiment was 25-30. They were graded from 0-3: no axons, 0; axons growing in a pole of the explant, 1; axons growing at more than one area or throughout the explants, 2; and greater axonal outgrowth (usually greater than 1/2 of the explant length) throughout the explants, 3.
Dissociated neuronal cultures with DRGs were established using a standard protocol (17). In brief, L3-L5 DRGs from adult rats were dissected and transferred into Ca2+- and Mg2+-free Hank's buffered salt solution (CMF-HBSS) containing gentamycin (10 µg/mL). Enzymatic digestion of the DRGs was initiated by the addition of 0.125% collagenase type I (Sigma, St. Louis, MO, U.S.A.) diluted in CMF-HBSS, and incubating them at 37℃. After 30 min, the DRGs were incubated in 0.25% trypsin for 25 min at 37℃. After enzymatic digestion, the cells were dissociated by mechanical trituration through a fire-polished glass pipette. The cells (2000 per well) were plated into one well per eight-well plate (Labtek, Nunc, Rochester, NY, U.S.A.) coated with poly-l-lysine (500 µg/mL), and cultured for 36-40 hr in the presence or absence of netrin-1. The neurites were quantified after immunostaining of the neurons with an anti-β-tubulin antibody. A cell with neurites longer than the cell body diameter was defined as a neurite bearing cell, and the number of such cells was counted. The length of the longest neurite in a group of randomly selected 180-200 neurons was measured using the LSM 510 image analysis program (Carl Zeiss). The neurite lengths between the groups were compared using the Student's t-test.
Reverse transcription-polymerase chain reaction (RT-PCR)
The mRNA expression of dcc and unc5h1-4 was examined by RT-PCR. The total RNA was isolated from adult spinal cords and DRGs by using Trizol reagent (Invitrogen), according to the manufacturer's instruction. First-strand cDNA was synthesized in 20 µL reaction mixtures with 1 µg of the total RNA using the M-MuLV reverse transcriptase (New England BioLabs, Ipswich, MA, U.S.A.). PCR reaction was performed with the following primers. Primers for amplification and the size of PCR products are as follows: DCC forward; 5'-AAGAATGGAGATGTGGTGATTCC-3', DCC reverse; 5'-CCAGTTGTCACCCTCTCTGGAG-3'(size of the PCR product: 325 bp), unc5h1 forward; 5'-CACCACCACTACCACCTACCAGG-3', unc5h1 reverse; 5'-CCCGAGGAAGTTGAAGG TCCC-3'(241 bp), unc5h2 forward; 5'-CGACCCTAAAAGCCGCCCC-3', unc5h2 reverse; 5'-GGGATCTTGTCGGCAGAGTCC-3'(318 bp), unc5h3 forward; 5'-AGGCTG CTCCTGACTCAGATG-3', unc5h3 reverse; 5'-GGGTCTAGAATTGGAGAATTGG-3'(313 bp), unc5h4 forward; 5'-CATAGAGGAGCCTGAAGATG-3', unc5h4 reverse; 5'-ACGGAAGCTTTCCTACTCTT-3'(218 bp), rat GAPDH forward; 5'-TGCCGCCTGGAGAAACCTGC-3', rat GAPDH reverse; 5'-TGAGAGCAATGCCAGC CCCA-3'(172 bp). The PCR cycling consisted of an initial 5 min at 94℃, followed by (30-32) cycles of 60 sec at 94℃, 45 sec at 54-62℃, and 60 sec at 72℃, with a final incubation of 5 min at 72℃. The PCR products were visualized by ethidium bromide staining (1 µg/mL), and the relative intensity (RI) index of the DNA bands from three independent experiments was analyzed using a luminescent image analyzer (LAS-3000, Fujifilm, Tokyo, Japan). The statistics of PCR results were obtained using Student's t-test.
In situ hybridization
The generation of unc5h1 and unc5h2 cRNA probes was performed using an in vitro transcription kit with digoxigenin-11-UTP (Roche Diagnostics, Nutley, NJ, U.S.A.). The plasmids containing the fragments of the unc5h1 and unc5h2 which had been generated by RT-PCR were used as templates for the in vitro transcription. For in situ hybridization, frozen DRG sections from adult Sprague-Dawley rats (5 rats) were cut into 18 µm thickness by using a cryostat (Frygocut, Leica, Gallen, Switzerland). The sections were then fixed in 4% PFA for 10 min, washed three times with phosphate-buffered saline (PBS), and finally acetylated for 10 min. After prehybridization, the sections were incubated with the hybridization buffer (50% formamide, 4×SSC, 0.1% CHAPS, 5 mM EDTA, 0.1% Tween-20, 1.25×Denhartdt's solution, 125 µg/mL yeast tRNA, 50 µg/mL heparin, and 200 ng of digoxigenin-labeled probes) for 18 hr at 57℃. The amount and size of the transcribed cRNA were estimated with RNA gel electrophoresis. Non-specific hybridization was removed by washing in 2×saline-sodium citrate buffer (SSC) for 10 min at room temperature and the treatment with RNase A (10 µg/mL), followed by final washing with 0.1×SSC at 57℃ for 15 min. For immunological detection of digoxigenin-labeled hybrids, the sections were incubated with anti-digoxigenin alkaline phosphatase (Roche Diagnostics, 1:1,500) for 1 hr, and the color reaction was developed using 4-nitroblue tetrazolium chloride (330 µg/mL) and 5-bromo-4-chloro-3-indolyl phosphate (175 µg/mL). Sections were dehydrated and mounted with Crystalmount.
RESULTS
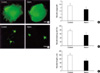
We examined whether netrin-1 regulates axon outgrowth of adult DRG neurons using three dimensional cultures and dissociated neuronal cultures. For three dimensional explant cultures, we cultivated lumbar DRGs from adult rats for 36-48 hr in Matrigel. We observed extensive growth of Tuj1-positive neurites in explants grown in Matrigel (Fig. 1A). The purified protein of netrin-1 (500 ng/mL) was sufficient to significantly block the regenerative neurite outgrowth from the explants. We scored the neurite outgrowth as described in Material and Methods, and found that the difference was significant (2.33±0.32 vs. 1.75±0.21, p<0.05). We also analyzed the effect of netrin-1 on the neurite outgrowth of adult DRG neurons in dissociated cultures. Tuj1 expression in cultured neurons was confirmed by immunostaining with an anti-Tuj1 antibody, and Fig. 1D shows that the number of Tuj1 positive neurons that have neurites (defined as a process longer than the cell body diameter) decreased from 76.84±6.2% (control) to 53.9±6.2% (netrin-1; p<0.05). We also examined the distribution of neurite length of the longest neurite within neurite-bearing cells, and found that the neurite length was also significantly decreased by netrin-1 treatment (Fig. 1C, E), when compared with the control. Thus, netrin-1 decreased both the number of process-bearing cells as well as the lengths of the neurites of the cells in acutely dissociated neurons. This finding indicates that netrin might inhibit the neurite formation and elongation in vitro. Since the intracellular cAMP level is known to modulate the actions of netrin in some neurons, we attempted to investigate the possible effect of db-cAMP and forskolin on the inhibitory function of netrin in the adult sensory neuron. However, we could not observe any significant effects (data not shown).
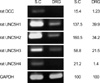
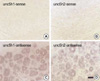
To identify netrin receptor genes that are expressed in the adult DRG, we performed semi-quantitative RT-PCR. We then compared the gene expression patterns of netrin receptors in the adult DRG with the patterns in the adult spinal cord, that have already been revealed in a previous study (12). This experiment showed that unc5h1-3 mRNAs, but not dcc mRNA, are expressed in the adult DRG (Fig. 2). The low level of dcc mRNA expression was observed only in the adult spinal cord. Among unc5hs, the unc5h1 and unc5h2 showed a relatively high level of mRNA messages in the adult DRG. The expression of unc5h4 mRNA was not detected in adult DRG, whereas a low level of unc5h4 mRNA expression was found in the spinal cord. In order to understand the cellular localization of the unc5h mRNAs, we examined the mRNA expression using the in situ hybridization method (Fig. 3). Because unc5h1 and unc5h2 are major genes among the Unc5 family that are expressed in the adult DRG, we performed in situ hybridization histochemistry with unc5h1 and unc5h2 antisense cRNA probes. Consistent with the result of the RT-PCR, this experiment clearly showed that unc5h1 and unc5h2 are abundantly expressed in the adult DRG. Two mRNA messages were primarily localized in sensory neurons regardless of the cell size, even though we could not completely exclude the mRNA expression in the non-neuronal cells in the DRG. However, the sense probes did not label any positive structures in the adult DRG sections. Together, these data indicate that Unc5h receptors are expressed not only in the adult spinal cord but also in the adult DRG and that Unc5h receptors may be involved in the inhibition of neurite outgrowth from adult DRG neurons via netrin-1.
DISCUSSION
In the present study, we found that netrin treatment inhibits the regenerative axon growth of adult DRG neurons. It has been reported that netrin may play a role in peripheral nerve regeneration (14, 18-20). Madison et al. reported that netrin expression in the adult sciatic nerve is dramatically increased after sciatic nerve injury (19). According to this report, netrin induction was found in Schwann cells. The glial origin of netrin was also demonstrated in the adult optic nerve (15). Therefore, netrin secreted from Schwann cells or other glial cells in injured nerves may affect the regeneration of peripheral nerves either directly by acting on neurons or indirectly by regulating the glial cell behavior. Our data that show the expression of unc5h1-3 mRNA in the sensory neurons of the adult DRG may support the idea that netrin, present in the sciatic nerve, regulates nerve regeneration directly through the receptors expressed in the peripheral sensory neurons. This hypothesis appears to be consistent with the finding of Watanabe et al. (21), which revealed the expression of unc5h mRNA in the embryonic DRG neurons and the inhibition of neurite outgrowth from the embryonic DRG by netrin treatment.
It was recently reported that netrin-1 in the dorsal part of the embryonic spinal cord acts as an inhibitory cue for primary sensory axons during development, thereby facilitating appropriate formation of sensory neural networks (21). However, the participation of netrin in spinal cord regeneration remains uncertain. In the adult spinal cord, netrin expression is known to be increased after the spinal cord injury, and inflammatory cells in the wound region are responsible for the high levels of netrin mRNA expression after the injury (14). In the present study, we demonstrated that netrin-1 inhibits neurite outgrowth from adult DRG neurons. Thus, our data put forth an interesting hypothesis that netrin in the wounded area of the spinal cord may inhibit or repel the regenerative axonal growth from DRG neurons into the dorsal column after spinal cord injury in adult rats. Alternatively, netrin may be involved in the axonal retraction after the injury. In this sense, netrin could be a novel inhibitory molecule for spinal cord regeneration.
The dominant expression of Unc5h as compared to DCC expression in the adult DRG is similar to the expression pattern of Unc5h in the adult spinal cord. Therefore, the expression of the attractive netrin receptor, DCC, appears to be downregulated either by the turn-off of gene transcription (23) or by the ubiquitin-mediated degradation during development (17). On the other hand, Unc5h expression continues and even increases with postnatal neuronal maturation in the DRG. According to the previous studies, unc5h1 and unc5h3 mRNA expression was observed in the DRG during embryonic development (21, 24), and a mutation in unc5h3 resulted in the aberrant projection of DRG axons. Because Unc5h, the receptor mediating the repulsive function of netrin, is mainly found in the adult DRG and spinal cord neurons, netrin in the adult neurons may be a neurite-inhibitory molecule. Thus, it could be possible that the inhibitory response of adult DRG neurons to netrin is attributable to Unc5h receptors. However, further studies are required to elucidate the in vivo function of netrin/Unc5h signaling in the adult nervous system.
 XML Download
XML Download