

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Yeasts are typically opportunistic pathogens in the human oral cavity and have been detected in 7 - 18% of infected root canals.1 One of the predominant fungal pathogens in oral and vaginal infections is Candida albicans (C. albicans).2 They are considered to be dentinophilic microbes because of their capacity to colonize dentinal walls and invade dentinal tubules.3 Additionally, C. albicans can grow as a biofilm4 that is over 100 fold resistant to antifungal agents such as fluconazole.5 The resistance of these biofilms is due to reduced rates of metabolism and growth, the presence of an exopolysaccharide matrix and protective factors, and their stress response.6 Indeed, C. albicans have been found in therapy-resistant endodontic infections.78
Mechanical instrumentation alone cannot completely remove microorganisms from the root canal system.9 Therefore, an intracanal medicament is recommended in order to prevent multiplication and recovery of residual microorganisms.10 The most commonly used intracanal medicament, calcium hydroxide (CH), is available to reduce microbial remnant.11 However, C. albicans showed resistance to aqueous CH.12 Nystatin (Nys) is one of the efficient antifungal agents against C. albicans.13 Nonetheless, the antifungal resistance of Nys increased when the drug was repeatedly used.14
Antimicrobial peptides (AMPs) have a wide spectrum of antimicrobial activity against oral microorganisms, and there was a study about the antimicrobial peptide-based treatment for endodontic infections using several AMPs that are endogenous to the oral cavity.15 These include human β-defensins (HBDs), which are cationic peptides expressed in inflamed dental pulps.16 HBDs (HBD1, 2, 3, and 4) have antimicrobial properties, and HBD3 was found to be the most potent.17 HBD3 possessed a broad spectrum of antimicrobial activities against Gram-negative, Gram-positive bacteria, and fungi with strong immunoregulatory activity.18
Although we recently demonstrated the broad antimicrobial effects of the synthetic peptide consisting of only 15 amino acids from the C-terminus of HBD3 (HBD3-C15, GKCSTRGRKCCRRKK) against endodontic bacteria in either single species (Enterococcus faecais) biofilm19 or multispecies biofilms in vitro,20 its antifungal properties are poorly understood. A previous study demonstrated a peptide consisting of 22 amino acid from the C-terminus of HBD3 (HBD3-C22) was effective against C. albicans.21 Therefore, the purpose of this ex vivo study was to compare the antifungal activity of a synthetic peptide consisting of 15 amino acids at the C-terminus of human β-defensin 3 (HBD3-C15) with CH and Nys against C. albicans biofilm.
Materials and Methods
Effects of HBD3-C15 on C. albicans biofilm on dentin disks
This study was approved by the Institutional Review Board of Seoul National University Dental Hospital, Seoul, Korea (CRI 15007). Single-rooted premolars with fully formed apices (n = 10) were collected from patients undergoing extractions for orthodontics in the Department of Oral and Maxillofacial Surgery at Seoul National University Dental Hospital. Their root surfaces were cleaned of calculus and soft tissue with an ultrasonic scaler, and stored in 0.5% sodium azide (Sigma-Aldrich, St. Louis, MO, USA) at 4℃. The roots were sliced into 500 µm thick cross sections using an Isomet precision saw (Buehler, Lake Bluff, IL, USA). These dentin disks were treated with 17% ethylenediaminetetraacetic acid (pH 7.2, Sigma-Aldrich) for 5 minutes, followed by 2.5% sodium hypochlorite (Sigma-Aldrich) for 5 minutes. Disks were then neutralized with 5% sodium thiosulfate (Sigma-Aldrich) for 5 minutes, and finally washed three times with distilled water. They were autoclaved for 15 minutes at 121℃, and incubated in liquid growth medium containing peptone-yeast-glucose (PYG) in 10 mmol/L potassium phosphate-buffered saline (pH 7.5) at 37℃ for 24 hours to confirm sterility. C. albicans (KCTC 7270, Korean Collection for Type Cultures, Daejeon, Korea) were grown in yeast malt media at 37°C until they reached mid-log phase (A600 = 0.1). Replicate dentin disks were incubated with cell aliquots (300 µL/well, 6 × 106 cells/mL) in 48-well plates for 48 hours and treated with either HBD3-C15 (0, 12.5, 25, 50, 100, 150, 200, or 300 µg/mL, NIBEC, Seoul, Korea), aqueous CH (100 µg/mL, DC chemical Co. Ltd., Seoul, Korea), Nys (20 µg/mL, Sigma-Aldrich), and saline (control) group for 7 days at 37℃. HBD3-C15 peptide was prepared by F-moc-base chemical solid-phase, serially diluted in sterile distilled water, and then applied on dentin disks. Subsequently their surfaces were examined by field-emission scanning electron microscopy (FE-SEM, S-4700, Hitachi, Tokyo, Japan).
LIVE/DEAD Biofilm viability assay
C. albicans mid-log phase cultures (3 mL/dish, 6 × 106 cells/mL) were transferred to a cover glass bottom dish (SPL lifescience, Pocheon, Korea) and incubated for 48 hours before being treated with either HBD3-C15 (0, 12.5, 25, 50, 100, 150, 200, and 300 µg/mL), aqueous CH (100 µg/mL) or Nys (20 µg/mL) triplicate dishes of each for 7 days at 37℃. Each dish was aspirated to remove broth, and washed gently with PBS. Finally they were stained with the FilmTracer LIVE/DEAD Biofilm viability kit (Molecular Probes, Carlsbad, CA, USA), which uses SYTO9 and propidium iodide (PI) to stain live and dead cells within biofilms respectively. SYTO9 stains both live and dead microorganisms fluorescent-green, whereas PI stains only the nucleic acids of cells with damaged membranes, and thereby identifies dead microbes. The stained C. albicans biofilms were examined by confocal laser scanning microscopy (CLSM, LSM 700, Carl Zeiss, Jena, Germany) with the ×40 lens. CLSM images were acquired by using ZEN 2010 (Carl Zeiss) software at a resolution of 512 × 512 pixels with a zoom factor of 2.0. Each 2 dimensional (2D) image covered an area of 230.34 × 230.34 µm. The 3 dimensional (3D) reconstructed images had a z step of 1 µm in each stack, and there were 15 stacks in total.
Image and statistical analyses
The CLSM images were analyzed with bioImage L (http://bioimagel.com) software. The green and red stained portions of the biofilm were used to calculate live and dead cell subpopulations within the total biomass. Statistical significance was examined using a two-tailed t-test, a one way analysis variance, and a post hoc test at a significance level of p = 0.05 using SPSS ver. 22 (SPSS Inc., Chicago, IL, USA).
Results
HBD3-C15 and Nystatin showed antifungal activity against C. albicans on human dentin
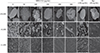
The biofilm of C. albicans on dentin disks was inhibited by HBD3-C15 in a dose-dependent manner (Figure 1). Comparing with CH group, there were fewer C. albicans aggregated on dentin in the groups of HBD3-C15 (≥ 100 µg/mL) and Nys (Figure 1). Ruptured cells were also observed at the concentrations of 150 to 300 µg/mL of HBD3-C15 and Nys (Figure 1).
HBD3-C15 and Nystatin reduced C. albicans survival and biofilm
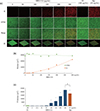
CLSM demonstrated that C. albicans survival and biofilm were reduced by HBD3-C15 in a dose-dependent manner (Figures 2a and 2b). The dead cell biomass in the HBD3-C15 (≥ 100 µg/mL) and Nys was significantly more (p < 0.05) than that of CH group (Figure 2c), but not significantly (p < 0.05) different between HBD3-C15 (300 µg/mL) and Nys.
Discussion
Native (whole) HBD3 is a 45-amino-acid protein that acts as a cationic antimicrobial agent.22 A previous study demonstrated that the whole recombinant HBD3 peptide had antifungal activity on C. albicans.23 However, the stability of the whole HBD3 peptide is weak and its half-life is short.19 Therefore, we focused on the most cationic residues at the C-terminus of HBD3 and created an even shorter synthetic peptide HBD3-C15, which would have the potential for greater antifungal activity at a reduced cost. As expected, the synthetic HBD3-C15 peptide showed superior antifungal activity than CH, and the effect was increased in a dose-dependent manner. Another study also reported that synthetic HBD3-C22 (SCLPKEEQIGKSTRGRKCRRKK) peptide had antifungal activity against C. albicans, while its activity was less salt sensitive and its synthesis was more affordable than the whole HBD3.21
Dentin disk consists of a hydroxyapatite containing collagen matrix,24 and biofilms grown on hydroxyapatite have been shown to exhibit considerable adherence and resistance to displacement.25 In the present study, FE-SEM showed that C. albicans aggregated on the dentin disks and the aggregation became fewer in the groups of Nys and HBD3-C15 in a dose dependent manner. CH showed limited efficacy in killing C. albicans in the form of biofilm, which is consistent with previous reports.2627 This could be due to the buffering capacity of the dentin against dissociated hydroxyl ion.28 Furthermore, C. albicans could survive at a wide range of pH values, the alkalinity of CH solution might not have antifungal effect on C. albicans.27 In addition, CH could provide the Ca2+ ions necessary for the growth and morphogenesis of Candida.27
Nys is polyene antifungal antibiotics, and its fungicidal effect by an alteration of cellular permeability causes loss of essential components from the cell,29 thereby leading to the cell death.30 Nonetheless, the administration of antibiotics into the root canal system is with the potential risk of adverse systemic effects, particularly toxicity, allergic reactions, and development of resistant strains of microbes.31 The present study also evaluated the inhibitory effects of the HBD3-C15 on C. albicans biofilm on a cover glass bottom dish and the percentage of dead cells in the entire biomass was a significantly increased at the concentration of 100 µg/mL more over, which we suggest that the minimal antifungal concentration of HBD3-C15 may be 100 µg/mL against C. albicans biofilm.
The antifungal mechanism of HBD3 against C. albicans involve specific interactions with the cell-surface proteins Ssa,32 which causes cell membrane disruptions.33 In the present study, ruptured C. albicans cells were frequently observed in the high concentrations of HBD3-C15. This suggests that HBD3-C15 increased the permeability of fungal cell membranes, thereby enhancing their uptake of PI, which is a small molecule (668.39 Da) that cannot cross intact membranes. However, the details of this killing mechanism are still not understood. Further studies will be needed to fully elucidate the antifungal pathways of HBD3-C15 peptide at the molecular level.
Synthetic HBD3 peptide has many advantages when it is used as an intracanal medicament in endodontic fields. Its antimicrobial efficacy and range of target microorganisms could be modulated by designing analogs of HBD3.18 HBD3 has a very low cytotoxicity against host cells.34 Furthermore, HBD3 can create synergy when incorporated with other antimicrobials and disinfectants such as chlorhexidine.35 In addition, an HBD3 containing gel will have a low viscosity that will prolong the contact with root canal walls and dentinal tubules and promote delivery to inaccessible anatomical structures such as fins and isthmuses, or curved canals by injection. The present result also showed that synthetic HBD3 peptide has a potential as a medicament in C. albicans-infected root canal. Together with the present anti-fungal effect and the previous antimicrobial effects against single-species (E. faecalis) biofilm19 or multi-species biofilm,20 HBD3-C15 peptide may be applied as an injectable intracanal medicament for therapy-resistant/persistent root canal infection or for endodontic regenerative procedure of infected immature permanent tooth.
 XML Download
XML Download