

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Hair graying is a common process occurring in people as they age. Fifty percent of the population has about 50% gray hair at the age of 50 years, known as the 50-50-50 rule12. Most non-pigmented hairs are white, attributable to total loss of melanin in the hair bulb. The term ‘gray hair’ refers to an admixture of white non-pigmented hairs and pigmented hairs. Sometimes a single hair fiber can show a progressive dilution of pigment from black, through gray to white. Hair graying may be inherited in an autosomal-dominant manner34. Hair graying usually develops initially at the temples, and then spreads to the frontal area, the vertex, and the parietal region, affecting the occipital region last5. If graying occurs prior to the third decade of life, this is generally termed premature graying, although the criteria may differ56. Hair graying is associated with various autoimmune disorders including vitiligo and several rare premature aging syndromes including the Hutchinson Gilford and Werner'syndrome7. Hair graying may also reflect nutritional deficiencies (especially of vitamins) and the use of drugs such as chloroquine8910. We do not know exactly the cause of hair graying in humans. Some evidences were found in mouse, but still only partial. Here, we reviewed previous studies on the mechanism of graying in humans and seek to offer some new insights.
INVOLVEMENT OF BULBAR MELANOCYTES IN HAIR GRAYING
Tobin and Paus5 proposed that graying is caused by the depletion of hair follicle bulbar melanocytes, due to dysregulation of antioxidant mechanisms and expression of anti-apoptotic factors. Overproduction of copper-zinc superoxidase induces excessive H2O2 formation and triggers oxidative damage. Accumulated reactive oxygen species (ROS) imposes significant oxidative stresses on both bulbar melanocytes per se and the highly proliferative hair bulb epithelium (consisting of keratinocytes)11. Inflammation, ultraviolet light, cigarette smoking, psychoemotional stress, certain chemicals, and genetic defects may trigger oxidative stress, inducing graying121314. It was suggested that graying may be caused by reduced tyrosinase activity in bulbar melanocytes or an abnormal interaction, melanosomes transfer, between these melanocytes and the cortical keratinocytes of hair bulbs1215. Also, graying may result from insufficient melanocyte migration from the reservoir (the hair bulge) of the upper outer root sheath (ORS) to the hair bulb lying closest to the dermal papilla5. The levels of both anti-apoptotic factors (including BCL-2) and melanogenic enzymes (TRP-1 and TRP-2) are valuable markers of graying. The mitochondrion is the primary target of oxidative stressors, compromising energy production. Thus, a mitochondrial theory of aging was suggested to explain hair graying13.
Melanin transfer from melanocytes to cortical keratinocytes may reduce keratinocyte proliferation and increase terminal keratinocyte differentiation, probably by modulating intracellular calcium levels1617. The growth rate of non-pigmented (white) beard hair is greater than that of adjacent pigmented hair18. Melanin granules transferred to keratinocytes may serve as regulatory packages, controlling cell differentiation and metabolic status1920. Together, the data suggest that non-pigmented (white) hairs may grow more rapidly and become thicker than pigmented hairs, due to an absence of melanin in bulbar keratinocytes. Active hair growth of white hairs may be considered as a result, not cause, of hair graying.
Insufficient neuroendocrine control of hair follicle melanogenesis by locally synthesized factors including adrenocorticotrophic hormone, thyrotrophin-releasing hormone, thyroid hormone, α-melanocyte stimulating hormone, and β-endorphin, melanocortin has been also suggested to trigger or progress graying2122232425262728. Clinical interventions seek to modulate neuroendocrine factor-mediated control of melanogenesis and human melanocyte biology, to prevent or reverse graying29.
Interestingly, graying is sometimes reversed even in the elderly. For example, some non-pigmented (white) hairs begin to produce melanin once more after exposure to radiation or cytokines, becoming proximally pigmented. One explanation is that melanocytes negative for 3,4-dihydroxyphenylalanine and most melanocyte-specific makers remain in the ORSs of white hairs and begin to produce pigment after stimulation3031.
Thus, European researchers, including Paus and Tobin, considered that graying is caused by depletion or dysfunction of melanin-producing melanocytes located in the hair matrix near the dermal papillae of hair follicles. However, amelanogenic melanocytes are also present in the ORSs. Such melanocytes may be able to migrate and differentiate into melanogenic melanocytes, when certain stimuli are applied. Many researchers have sought to identify such re-activating molecules recently.
INVOLVEMENT OF BULGE MELANOCYTE STEM CELLS IN GRAYING
Nishimura et al. focused on the role played by melanocyte stem cells (MSCs) in the hair bulge, as opposed to follicular melanocytes, reporting that graying is caused by defective MSC self-maintenance. Such maintenance requires BCL-2 and the micro-ophthalmia-associated transcriptional factor (MITF). Also, transforming growth factor-β (TGF-β) and collagen XVII (Col17A1) are involved in regulation of MSC maintenance323334.
Col17A1, a hemidesmosomal transmembrane collagen, is highly expressed in follicular stem cells. MSCs of the hair bulge do not express Col17A1, but premature graying and hair loss were evident in mice lacking Col17a1. Expression of COL17A1 in basal keratinoctyes and follicular stem cells of COL17a1-deficient mice blocked premature MSC differentiation and restored TGF-β signaling. Hair follicular stem cells in the bulge-sub-bulge area provide a functional niche for the MSCs35.
Defective MSC self-maintenance attributable to a Bcl-2 deficiency triggers selective MSC apoptosis, but not apoptosis of bulbar melanocytes. Aging or injury to MSCs induces ectopic pigmentation and/or differentiation within the hair bulge niche, accelerated by Mitf, a key transcriptional regulator of melanogenesis. The roles played by stem cell apoptosis and ectopic differentiation remain unclear; both may contribute to MSC loss to a similar extent32. However, the work of Fisher and Nishimura is essentially mouse-based. Further study should be done in humans to unveil hair graying.
Graying is associated with age-related defects in MSC maintenance. Irreversible DNA damage caused by genotoxic stressors such as ionizing radiation interrupts MSC comeback in mice. MSC depletion via differentiation or apoptosis triggers ‘irreversible’ graying. A defect in the ataxia-telangiectasia mutated (ATM) gene, a kinase serving to transduce the DNA damage response, induces ectopic MSC differentiation. ATM action is important for the maintenance of MSC stemness36.
Recent work has focused principally on MSCs of the hair bulge, not on bulbar melanocytes. The B-Raf and C-Raf kinases that play critical roles in melanoma development are required for MSC maintenance, but not for melanocyte lineage development. Hair graying was evident in a double-Raf knockout mouse37. Human amelanotic melanocytes may be considered to be analogous to the MSCs of mice. The the molecules including BCL-2, MITF, B-Raf, B-Raf, TGF-β, Pax3, Wnt, and Notch have been suggested to be useful markers of human MSCs. However, no specific marker of MSCs has yet been identified, because it is difficult to perform genetic studies in humans38.
We found that Sox10, Pax3, and MITF-M (specific markers of MSCs or melanocytes) were absent from the bulbs of white, but not black, hairs in humans. Thus, both melanocytes and MSCs or amelanotic melanocytes were absent from the bulbs, suggesting that graying may be caused by a depletion of MSC migrating into the hair bulb39.
ACTIVE HAIR GROWTH AND GRAYING
Earlier, we presented data supporting the idea that graying was attributable to MSC depletion in the hair bulge39. We found the difference in genes/proteins expression associated with thickness and mechanical property, e.g., keratins (KRTs) and keratin-associated proteins (KRTAP), between pigmented and non-pigmented (white) hairs. Interestingly, Van Neste40 found that non-pigmented (white) hair follicles grow significantly more rapidly than pigmented hair follicles in organ culture. Clinically, the growth and thickness of non-pigmented follicles are significantly greater than those of pigmented follicles41.
Our previous studies showed that a few long hairs on the eyebrow are white, while short hairs on the eyebrows are usually black41. There is no report that hair graying may be more severe in farmers than office workers. These may suggest that external stress such as ultraviolet light may be less important in terms of graying than was formerly thought. Thus, we propose that graying is attributable principally to active hair growth.
As expected, we found that KRTs, including KRT6, KRT14/16, and KRT25, were expressed more extensively in non-pigmented (white) than pigmented (black) hairs. Many isoforms of KRTAP were upregulated in white compared with black hairs4243. The expression level of fibroblast growth factor 5 (FGF5), an upstream inhibitor of KRT and KRTAP synthesis, was reduced in white compared with black hairs4445. In contrast, the expression level of FGF7, an upstream stimulator of KRT and KRPAP synthesis, was increased in white compared with black hairs4647. In general, genes and proteins associated with active hair growth are expressed more prominently in white (non-pigmented) than black (pigmented) hairs. Active hair growth, combination of thickness and rate of growth, may result from upregulation of metabolic activity and cell divisions, which may lead to ROS production and MSC depletion in hair follicles. Thus, we suggest that graying is associated with active hair growth41; however, confirmation is required.
As discussed above, non-pigmented (white) hairs grow more rapidly to form thicker bundles, than do pigmented hairs, as a consequence of the absence of melanin in bulbar keratinocytes. Many researchers believe that the active growth of non-pigmented (white) hairs simply reflects the absence of melanin in such hairs. However, there is no evidence that non-pigmented hairs grow more rapidly or thickly than pigmented hairs in vivo.
Animal models of active hair growth such as the FGF5-deficient mouse48, which has longer body hairs than the wild-type mouse, may prove our theory. If we are correct, gray hairs may appear earlier in aging FGF5-deficient mice than in age-matched wild-type mice. Clinically, long-term follow-up of changes in the longer and thicker ‘pigmented’ hairs of the eyebrow, which eventually become non-pigmented (white), may also validate our theory.
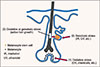
Together, our data suggest that graying may be attributable to active hair growth. Thus, graying may be delayed by suppressing such growth. This could be much more effective than seeking to revert white (non-pigmented) hairs to pigmented hairs, as white hairs containing MSCs or amelanogenic melanocytes are rare in the human scalp. The summary diagram for three theories for the mechanism of hair graying is shown in Fig.1.
CONCLUSION AND PERSPECTIVES
Hair graying is closely associated with MSC depletion in the hair bulge. Although re-activation of remaining MSCs or amelanogenic melanocytes may sometimes restore pigmentation, hair bulb melanocyte dysfunction may contribute temporarily to graying. The questions of whether melanin in hair follicles suppress hair growth and whether the principal cause of MSC depletion during graying is external (ultraviolet light, etc.) or internal (active hair growth), remain open.
 XML Download
XML Download