

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In 1981, Doll and Peto published the results of an epidemiology study to estimate the causes of cancer in the United States [1]. The conclusion of that study was that between 1/3 and 2/3 of all human cancers are preventable. Certainly smoking is the cause of about 30% of cancers but they also estimated that in addition to stopping smoking, dietary change was a major strategy for preventing cancer [1].
The important influence of diet on carcinogenesis is further supported by the more recent literature surveys sponsored by the American Institute for Cancer Research (AICR)/World Cancer Research Fund [2,3]. These reports were produced by a team of distinguished scientists who surveyed the existing literature, including both basic science and epidemiology reports, on diet and cancer. A conclusion of the first report [2], released in 1997, was that 'inappropriate diets cause about one-third of all cancer deaths' (p.7, [2]) and it was estimated that 30% to 40% of cancer could be prevented by a recommended diet change, physical activity and appropriate body mass (p.7, [2]).
During the next 10 years, the scientific literature reporting the results of research on diet and cancer dramatically increased, thus, a committee was commissioned to update the 1997 AICR report. These findings, published in 2007 [3], again support the notion that cancer is largely a preventable disease. In addition, the scientists compiling this report [3] chose to assess the effects of diet and lifestyle choices on other chronic diseases, including Type II diabetes and cardiovascular diseases, thus, making a recommendation for overall 'health promotion'. It was reported that a plant based diet coupled with exercise and maintenance of a healthy weight could reduce risk for all of these chronic diseases [3]. What common mechanism contributes to these common, chronic diseases that can be addressed by diet and lifestyle changes?
Inflammation increases risk for cancer and other chronic diseases
The link between inflammation and cancer was first noted as an association between inflammatory diseases and increased risk for cancer of that organ. For example, inflammatory bowel disease increases the risk for colon cancer [4], human papilloma virus increases risk for cervical cancer [5], schistosoma infection increases risk for bladder cancer [6] and Helicobacter pylori (and stomach ulcers) increase risk for stomach cancer [7]. The presence of chronic inflammation also contributes to the development of type 2 diabetes and cardiovascular disease [8, 9,10,11,12,13]. An underlying source of chronic inflammation can be obesity [13] effectively tying obesity into these chronic diseases. We now know that excess adipose tissue produces inflammation inducing cytokines [13], increases oxidative damage [9] and directly alters gene transcription [14]. All of these mechanisms can contribute to cancer development.
Chronic inflammation can have both positive [15] and negative [16] influences on tumor development. The cytokines produced by immune cells [such as macrophages, cytotoxic T cells (CTLs) and natural killer (NK) cells) can promote tumor immunity or promote the development of chronic inflammation [17]. However, the microenvironment of the tumor lacks normal immune-protective mechanisms and promotion of chronic inflammation dominates [17]. Cytokines found in the milieu of the inflammatory state can promote tumor cell growth [tumor necrosis factor α (TNFα), interleukin 6 (IL-6)], enhance tumor cell invasion [transforming growth factor β (TGFβ)], enhance metastasis (TNF α, IL-6, TGFβ), enhance angiogenesis (TNFα, IL-17, TGFβ) and impair the function of NK cells, CTLs and macrophages [17].
Inflammation and NFκB
Activation of the NFκB pathway provides a key molecular link between inflammation and carcinogenesis [18]. NFκB is a transcription factor that activates expression of multiple genes related to inflammation. It is also known as an 'oxidative stress responsive transcription factor' and is upregulated in response to oxidative stress. The mammalian NFκB family has five members, RelA (p65), c-Rel, RelB, p50 (NFKB1), and p52(NFKB2) [19]. The active form is a dimer of combinations of these five members. In the cytoplasm the dimer is found bound to IκB, (inhibitor of NFκB). On activation, the inhibitor is released and degraded in the proteosome, the dimer is phosphorylated and moves into the nucleus to bind DNA and to activate gene transcription [16,20]. Activation of NFκB provokes an autocrine, self-perpetuating response, that is, the products of many of the genes that are activated by NFκB (such as COX2 or TNFα) feedback and increase activation of NFκB [18,21].
Activation of NFκB is critical for normal immune response and for organogenesis [22], however NFκB has been found to be hyperactivated and to contribute to progression of multiple tumor types [16]. In organogenesis or in inflammatory responses the genes for cell survival, proliferation, angiogenesis and invasion that are activated by NFκB are critical for organ growth, development and wound healing [23]. However, when activated in a cancer, these same genes contribute to cancer growth and progression. Multiple carcinogens, inflammatory agents and factors in the tumor microenvironment have been found to activate NFκB [16]. Breaking the cycle of NFκB activation has been shown to slow tumor growth [24, 25,26]. Thus, one critical mechanism by which dietary components can suppress cancer growth and progression is by suppressing the activation of NFκB.
Dietary components that suppress NFκB and/or carcinogenesis
Dietary change is a key mechanism for reducing obesity and for providing components to suppress inflammation, decrease oxidative damage and change gene expression. It has been shown that at least part of the mechanism for suppression of carcinogenesis of many dietary components is by the suppressing the activation of NFκB. Some of these beneficial dietary components are reviewed below.
Fruits, vegetables, whole grains and nuts: Plants produce multiple nutraceuticals that have been shown to modify carcinogenesis via multiple pathways [27] thus increased consumption of plant products has been recommended to reduce risk for cancer [28,29].
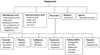
Polyphenols: Plants produce varying forms of polyphenols, a large, diverse class of compounds, many with antioxidant properties [30]. Structurally, polyphenols all have one or more aromatic (phenolic) rings with different structural elements that allows classification into subgroups [30]. In addition to (or perhaps because of) their antioxidant properties, most polyphenols have a common mechanism of decreasing the activation of NFκB. As such, a diet with generous amounts of fruit, vegetables, whole grains and nuts will provide multiple polyphenols for suppression of NFκB. See Fig. 1 for a diagram of polyphenol classes. Some especially well researched polyphenols are highlighted below.
Isoflavones
Isoflavones comprise a subclass of naturally occurring phenolic flavonoids and have a structure similar to estrogen. Isoflavones are produced almost exclusively by the members of the Fabaceae (i.e., Leguminosae, or bean) family. Soybeans (which contain high amounts of the isoflavone, genistein) are a common source of isoflavones in human food. Genistein can act as a phytoestrogen in mammals and, though they are not steroids, can alter activity of the estrogen receptor [28,30,31]. Some isoflavones are termed antioxidants because of their ability to trap singlet oxygen. Genistein has been shown to suppress the activity of NFκB when arachidonic acid (an omega 6 fatty acid which usually promotes inflammation) is included in the diet and to complement the activity of docosahexaenoic acid (DHA, which is usually anti-inflammatory) against breast cancer [32]. Diadzein, another isoflavone, has been shown to decrease proinflammatory cytokines and suppress chronic inflammation in obese mice [33].
Flavonols
The subclass flavonols are the most ubiquitous of flavonoids in food [30]. Flavonol synthesis is stimulated by light so there can be large differences in flavonol concentration between different pieces of the same fruit depending on light exposure [30]. Quercetin has been shown to have anticancer activity by multiple mechanisms including: induction of apoptosis [34], inhibition of COX2 activity [35], enhancing natural killer cell activity [36] and inhibiting NFκB [27]. Quercetin has also been shown to enhance the activity and to reduce the detrimental side effects of doxorubicin [37]. Kaempferol has been shown to promote antioxidant defense (which would help suppress activation of NFκB), to prevent cancer and to slow growth of existing cancer [38].
Flavanols
The flavanols include catechins and gallocatechins that are found in many types of fruit, in green tea, red wine and chocolate [30]. This class of compounds has been found to suppress proliferation of many cell types and to suppress activation of NFκB and the associated inflammation [39,40]. Epigallocatechin, found in green tea, is one of the best known of these compounds and has been found to suppress carcinogenesis by multiple mechanisms including suppression of NFκB activation [41,42,43].
Stilbenes
Resveratrol is found in red wine. Results of studies on cell culture and animal models have mostly shown that resveratrol slows cancer growth, reduces inflammation [44] and decreases the activation of NFκB [45]. However, due to the poor bioavailability of resveratrol, the applicability in human cancer prevention is uncertain. Studies are ongoing to find methods to increase the bioavailability of resveratrol. There is indication that the multiple polyphenols found in red wine (anthocyanins, flavanols, flavonols, catechins, resveratrol) can act synergistically to suppress carcinogenesis more than would be expected from the amounts of the individual components [46].
Hydrobenzoic Acids
Ellagitannins and gallotannins (hydrolysable tannins) are converted to ellagic acid in the upper gastrointestinal tract and then to urolithins by gut microflora in the lower tract [47]. The urolithins are then bioavailable and have potent antioxidant activity. Ellagitannins and gallotannins are primarily consumed in berries [48] and other fruits including pomegranate [49]. These tannins have been found to have multiple health benefits including cancer suppression [48,50,51].
Hydroxycinnamic acids
Curcumin, a polyphenolic derivative of hydroxycinnamic acid [52], has been shown to have benefit against multiple diseases, at least partly by protecting against oxidative stress and inflammatory damage [53,54]. The clinical use of curcumin is limited by its poor bioavailability thus there is an active search for curcumin analogues with better bioavailability [54]. One mechanism for the activity of curcumin is to suppress NFκB [53].
Terpenes
Terpenes are isoprenoids which form the building blocks for the synthesis of molecules such as retinoids, tocopherols and lycopene. Both retinoids and tocopherols are well known antioxidants but they also have specific anticancer activity.
Retinoids
Retinoids are a class of compounds related to carotenoids, the dark green and deep yellow pigments of fruits and vegetables. They mediate anticancer effects via the RAR (retinoic acid receptor) and RXR (retinoid X receptor) pathways [55]. These pathways regulate cell proliferation, differentiation, and apoptosis and are essential for normal cell differentiation [55]. Disruption of retinoid signaling has been linked to multiple human cancers [55]. In humans, the retinoid, fenretinide, has been used as 'differentiation therapy' in combination with tamoxifen to help prevent recurrence of breast cancer [56]. Plasma carotenoid levels have been inversely correlated with human breast cancer risk [57]. The potential toxicity of high levels of synthetic retinoids is not a problem when carotenoids are consumed naturally in fruits and vegetables.
Lycopenes
Lycopene is a carotenoid that has antioxidant activity but does not have vitamin A activity; it is responsible for the red color of fruits and vegetables including tomatoes, watermelon and pink grapefruit [58]. Some studies indicate that lycopene is better absorbed after cooking and in the presence of fat, i.e., lycopene in spaghetti sauce may be more bioavailable than lycopene in raw tomatoes [58]. Lycopene has been shown to have anticancer activity against breast cancers by a variety of mechanisms and with varied potency depending on the genetic mutations of the cancer [59,60]. Dietary lycopene has been associated with decreased risk of death from prostate cancer [61].
Tocopherols
Vitamin E is also a well-known, lipid soluble antioxidant and is represented by 8 different molecules - four different tocopherols: alpha, beta, gamma and delta-tocopherol and 4 different tocotrienols: alpha, beta, gamma and delta tocotrienols [62]. In addition to its antioxidant activity, tocopherols can directly modulate gene expression [63,64]. The major dietary form of Vitamin E is gamma-tocopherol whereas synthetic tocopherol is usually a mix of isoforms. Gamma-tocopherol has been found to be effective at slowing growth of multiple cancer types, whereas alpha-tocopherol (high in vitamin E supplements) is largely ineffective [65,66].
Sulphur containing compounds
Sulphur containing compounds include isothiocyanates [found in cruciferous vegetables (broccoli, cauliflower, cabbage)] and the organosulfur compounds found vegetables of the allium family (onion, garlic).
Isothiocyanates
The isothiocyanates impart the characteristic sharp taste of wasabi, mustard, watercress, broccoli and other cruciferous vegetables. The isothiocyanates include allyl-, benzyl- and-phenethyl-isothiocyanate and sulforaphane. These molecules have been found to induce cancer cell death and to increase sensitivity to chemotherapy in multiple models [67,68,69,70,71,72]. The identified mechanisms were varied and range from induction of free radical mediated oxidative stress in some systems [67,72] to being antioxidative in other systems [68,73]. However, whatever the identified mechanism, the isothiocyanates protected against cancer development.
Organosulfur compounds
The consumption of allium vegetables, especially garlic have been linked to reduced risk of multiple cancers [74] especially prostate cancer [75,76]. As for many of the other natural compounds, multiple mechanisms of action, including suppressing inflammation, suppressing activity of NfκB and altering cell signaling pathways have been identified for the organosulfur compounds [74,76].
Melatonin
Melatonin is a potent antioxidant produced naturally by humans in the pineal gland and it is also produced by plants. In humans, melatonin has a wide variety of endocrine, neural and immune functions [77]. Melatonin has been shown to be chemoprotective against prostate [78], pancreatic [79], breast [80] and colon [81] cancer by multiple mechanisms [82] including reducing oxidative stress and inflammation [81,83], inducing apoptosis [82] and inhibiting angiogenesis [79].
Omega 3 fatty acids
Omega 3 fatty acids are commonly found in fish and fish oils. However, the 18 carbon omega 3 fat, α-linolenic acid (ALA) is made by plants and can be found in many plant foods. Canola and flax oil are especially high in ALA with low amounts of the corresponding omega 6 oil, linoleic acid. Omega 3 fats, not only ALA but also the longer chain eicosapentaenoic acid (EPA, 20 carbons) and docosahexaenoic acid (DHA, 22 carbons) have been studied as interventions in the growth of many cancer types. Suppression of the activation of NFκB has been identified as one mechanism of action of omega 3 fatty acids against multiple cancers (examples: [84,85,86,87,88,89,90,91,92,93,94,95,96,97]).
Use of walnuts as an intervention for cancer
Walnuts (English walnuts or Juglans regia) have multiple ingredients that individually (as described above) have been shown to have benefit for cancer prevention or suppression of growth of existing cancer. These ingredients in a single serving (1 ounce, 28.5g of walnuts) include ([98] except as noted): the highest amount of omega 3 fatty acid of any nut (2.5 g), polyphenols mostly anthocyanidins (0.8 mg) and proanthocyanidin (about 19 mg) and other antioxidants including vitamin A (6 IU), melatonin (3.5 ng/g [99]), gamma-tocopherol (5.9 mg), β-carotene (3 µg) and ellagic acid monomers [100], phytosterols (18.3 mg/100 g) and dietary fiber (1.9 g). There had been few to no studies using walnuts as an intervention for cancer prior to our work [101], but, with this mix of ingredients, we thought walnuts might be a whole food that would show benefit against cancer.
Our first study was to assess the effects of including walnut in the diet on the growth of existing cancer. We used a nude mouse model in which MDA-MB 231 human breast cancer cells were implanted between the scapulae. After the tumors had grown to measurable size, the mice were divided into two groups. One group was placed on a diet containing the AIN-76 diet modified to contain 10% total fat and to include human equivalent of 2 ounces per day of walnuts, the other group continued on the AIN-76 diet containing 10% corn oil as the fat. Diets were formulated to be isocaloric and isonutrient (see [101] for details). All animal work received approval from the Marshall University Institutional Animal Care Use and Committee prior to initiation.
As shown in Fig. 2 the tumors of the mice that consumed walnut in the diet grew significantly slower than the tumors of the mice that did not consume walnut. We found that the slower tumor growth could be related to a significant decrease in cell proliferation (proliferating cell nuclear antigen positive cells) and a slight but not significant increase in apoptotic cells. The omega 3 content of the walnut diet was reflected in the significant decrease in arachidonic acid (omega 6 fatty acid) and significant increases in the omega 3 fatty acids, eicosapentaenoic and docosahexaenoic acids in the liver [101].
In our second study (Fig. 3), we used a transgenic mouse model that develops mammary gland cancers to determine if regular consumption of walnuts could help prevent mammary gland cancers (all details in [102]). This was a transgenerational study; we started half of the mother mice on the walnut diet then assessed mammary gland cancer development in the female offspring that were randomized to control or walnut containing diets. The diets were the same as described in the first study (above). The results indicated that consumption of walnut after weaning reduced the risk for mammary gland cancer, however if the mother also consumed walnut (as would happen in a population that routinely includes walnut in the diet) then there was significant suppression of tumor development in the pups. Analyses of gene expression and protein in mammary glands without tumor indicated that reduced tumor risk was associated with suppression of NFκB activity by the walnut containing diet. We did not identify a single component of walnut that was responsible for decreasing risk for cancer development but rather proposed that it was the combination of ingredients in the walnut that reduced mammary gland cancer growth and development in mice.
CONCLUSIONS
Epidemiology studies have indicated that there is an environmental influence to cancer development and that overall diet or specific dietary components can increase or decrease cancer risk. Cell culture and animal studies have shown suppression of carcinogenesis by multiple components found in the fruits, vegetables, whole grains and nuts that would be consumed as part of a plant based diet. It seems difficult for humans to consume any single component that has been shown to be beneficial at a level and continuously enough to suppress carcinogenesis. However, the effects of the individual dietary components can be summed and increased when people eat a varied, plant based diet. The work of many scientists has shown that the individual components tested can reduce the production of inflammatory cytokines in animal and cell culture models. Since chronic inflammation contributes to the development of cancer, cardiovascular disease and diabetes, consumption of a varied plant-based diet, as recommended by multiple public health agencies, could effectively reduce the incidence of cancer and other chronic diseases.


 XML Download
XML Download