

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Cervical cancer is the fourth most deadly women's cancer worldwide [1]. In 2012, about 527,600 new cervical cancer cases were reported and around 265,700 deaths were attributed to cervical cancer globally, with less developed countries bearing the major portion of the disease burden [2]. In the United States, about 12,990 new cases of cervical cancer will be diagnosed and roughly 4,120 women will die from cervical cancer each year [3].
Currently, surgical removal of cervical tumor through radical hysterectomy is the treatment of choice in early cervical cancer and concurrent chemoradiation (CCRT) is the preferred modality for the cure of locally advanced cancer [45]. Radiation has been shown to induce DNA damage, leading to the loss of cell recovery, arrest of cell cycle, and cell destruction [6]. Chemotherapy has been considered as a primary treatment of metastatic cancer (cervical cancer IVB) and alternative treatment for recurrent cervical cancer patients who cannot be effectively treated using radiation or surgery. Chemotherapeutic agents that have been used for the treatment of cervical cancer include platinum drugs [78], alkylating agents [9], and anti-neoplastic compounds [1011]. Recently, Gynecologic Oncology Group (GOG) added bevacizumab on the conventional chemotherapy in cervical cancer [1213]. Bevacizumab is a humanized monoclonal antibody against angiogenic molecule vascular endothelial growth factor-A, and clinical study has shown that bevacizumab treatment can significantly increase the survival of patients with late stage cervical cancer [12]. However, in case of recurrent, metastatic cervical cancer, chemotherapy and radiation have shown limited efficacy to remove tumor burden with increasing toxicity arising from repeated treatments [121415]. Thus, the introduction of new treatments that could drive down the disease burden of cervical cancer is highly valuable.
Human papillomavirus (HPV) has been identified as an essential etiology of wart, cervical intraepithelial neoplasia (CIN) and cervical cancer [16]. In addition, HPV has been indicated to be a human biologic carcinogen for five other types of cancers: penile, vaginal, vulvar, anal, and oropharynx including the base of the tongue and tonsils, with a high percentage of oropharyngeal cancer cases in the United States being attributed to HPV [1171819]. Until now, over 200 types of HPV have been identified and subcategorized into high risk and low risk group according to the degree of oncogenic capacity [20]. High risk types of HPV include 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68, 72, and 82 and low risk types include 6, 11, 40, 42, 43, 44, 54, 61, 70, 72, and 81, with the high risk types HPV being considered as carcinogenic [2122]. Among high risk HPVs, type 16 and 18 are most carcinogenic for the progression of cervical disease and induce over 70% of cervical cancer [23].
HPV belongs to the Papillomaviridae family with non-enveloped, circular, and double-stranded DNA genome of about 8,000 base pairs that is covered with capsid proteins [24]. The HPV genome encodes a total of eight early and late genes. Early genes encode six early proteins E1, E2, E4, E5, E6, and E7, while the late genes produce L1 and L2 (Fig. 1) [2022]. The E1 and E2 proteins regulate replication and transcription of HPV DNA; E4 assist the viral release through the disruption of the cytokeratin of upper epithelium; E5 is responsible for inducing growth factors in host cell; E6 degrades tumor suppressor p53; and E7 binds and inhibits tumor suppressor pRb. Regarding the late genes, L1 and L2 encode the major and minor structural proteins that form the capsid of the HPV, respectively [24].
Fig. 1
The structure of human papillomavirus (HPV) and its function. HPV genome is composed of 8k base pairs and divided into early and late gene. The early genome encodes E1, E2, E4, E5, E6, and E7 and the late genome encodes L1 and L2. Oncoprotein E6 and E7 degrade tumor suppressor p53 and pRb, respectively. L1 and L2 build the structure of HPV capsid protein. LCR, long control region.
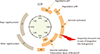
HPV infection in the basal cell layer of the cervical epithelium can be initiated through micro lesions of the tissues and cells [25]. There, HPV can multiply in an episomal state with its replication cycle being closely linked to the differentiation of the infected cell. In terminally differentiated cells, the HPV late genes produce capsid proteins to package virions to be released from superficial epithelial layers [26].
Alternatively, integration of HPV DNA into the genome of the infected cell may occur during persistent HPV infection [27]. The integration process often leads to the deletion of many early (E1, E2, E4, and E5) and late (L1 and L2) genes. Importantly, E2 is a negative regulator of the HPV oncogenes E6 and E7, and the absence of the E2 gene after integration leads to the elevated expression of these oncoproteins, which is thought to be an important process for the malignant change [1628].
The identification of HPV as an etiological factor for HPV-associated malignancies creates the opportunity for the control of these cancers through immunization and other target therapies. Currently, two types of preventive HPV vaccines, Gardasil (Merck, Kenilworth, NJ, USA) and Cervarix (GlaxoSmithKline, Rixensart, Belgium), are commercially available. The purpose of these vaccines is to block the entry of HPV into the epithelium of cervix by generating HPV-specific neutralizing antibodies against the major (L1) capsid proteins [2930]. However, although these preventive vaccines are effective in protecting against infection with oncogenic HPV types 16 and 18 [31], they cannot eliminate pre-existing HPV infections [32]. Consequently, there is a pressing need to develop effective treatments for established HPV infections and associated diseases. One potential treatment method involves the use of therapeutic vaccines that, unlike the preventive vaccines that intend to generate neutralizing antibodies against viral particles, aim to stimulate cell-mediated immune responses to specifically target and kill the infected cells. Since HPV oncoproteins E6 and E7 are required for the generation and maintenance of HPV-associated malignancies, they are consistently expressed in the transformed cells in HPV-induced cancer and precancerous lesions. Furthermore, because HPV E6 and E7 are foreign proteins, they can circumvent the issue of immune tolerance against self-antigens, a challenge presented by many other cancers, and thus they serve as ideal targets for therapeutic HPV vaccines [33]. These findings have inspired many attempts to create an optimal immunotherapeutic strategy against HPV infections and disease. In this review, we will cover various therapeutic vaccines in development for the treatment of HPV-associated lesions and cancers. Furthermore, we will discuss the potential of immune checkpoint inhibitors that have recently been adopted and incorporated into the design of immunotherapy against HPV-induced cervical cancer.
THERAPEUTIC HPV VACCINES
After HPV integrate into the genome of infected cells, HPV L1 and L2 genes are lost, and HPV L1 and L2 protein are no longer expressed in the infected cells [27]. As a result, the neutralizing antibodies induced by HPV preventive vaccine cannot effectively target these infected cells. Therapeutic vaccine aims to eliminate pre-existing HPV-associated disease by strengthen dendritic cell (DC) and antigen specific T lymphocyte in cellular immunity [34]. The tumor antigens, E6 and E7, are constitutively expressed only on the surface of cervical cancer and not on the normal tissue of cervix [3536], which can be the ideal target of therapeutic vaccine without damage to normal tissue.
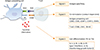
Antigen-presenting cells (APCs), such as the DCs, play a very crucial role in cellular immunity and determine the fate of naïve T cell. To make immune system active against cancer, APCs need to firstly uptake the foreign antigens released from tumor cells, process the antigens and present them to naïve CD8+ or CD4+ T cell using major histocompatibility complex (MHC) class I and II in the tumor draining lymph node, respectively. During antigen presentation, APCs induce several signals that drive the activation, proliferation, and differentiation of the T cells. The signal 1 is generated from interaction between T cell receptor and peptide-MHC complex which arise from antigen specificity and prepare the T cells for activation. After then, costimulatory receptor such as CD28 on T cell binds with costimulatory molecules CD80 (B7-1) and CD86 (B7-2) on APC and triggers survival and proliferation of activated T cells (Signal 2). Without the signal 2, immune tolerance and T cell anergy can be induced. Signal 3 arise from the orchestration of cytokines and guide the differentiation of T cell according to the environmental condition. Cytokines such as interleukin 12 (IL-12) and intercellular adhesion molecule 1 mediates the differentiation of naïve CD4+ T cells into T helper 1 (Th1) cell and cytokines including OX-40 ligand, IL-10 and transforming growth factor β (TGF-β) elicits TH2 cell polarization (Fig. 2) [37]. After activation, the matured and differentiated effector T cells migrate into tumor site and attack tumor cells.
Fig. 2
The mechanism of T cell stimulation and differentiation. T cell is stimulated through the complex of major histocompatibility complex T cell receptor (MHC-TCR) in the presence of antigen (signal 1). Co-stimulatory molecule is indispensible for the activation of T cell mainly through CD28 on T cell and B7 on antigen presenting cell (signal 2). Otherwise, T cell will fall into anergy state. Finally, T cell differentiation will be affected by cytokine or environmental factors (signal 3). ICAM, intercellular adhesion molecule; IL, interleukin; LFA-1, lymphocyte function-associated antigen-1; Th, T helper; OX-40, CD134.
This section discusses the various therapeutic HPV vaccines being developed and tested for their potential to elicit HPV-specific T cell-mediated immune response to clear HPV-associated diseases and cancers. These therapeutic vaccines include lie vector vaccines, peptide/protein vaccines, nucleic acid vaccines, and DC-based vaccines [38].
1. Live vectors
Live vector vaccine can provoke strong cellular and humoral immune response [39]. Live vector vaccines are categorized into two types, bacterial- and viral-based vector, according to the vector platforms. Live vectors can amplify target antigen during replications in the infected host cells. As a result, infection risk of live vector into host cell is evaluated carefully before treatment [40].
1) Bacterial-based vector
Lactobacillus casei, and Listeria monocytogenes (Lm) have been widely selected for the development of therapeutic HPV vaccines [41]. L. casei based vaccine is relatively safe and can potentially be administered orally. L. casei based vaccine expressing HPV E7 protein was given to patients with CIN III and E7 specific cellular immunity was evaluated with enzyme-linked immunospot (ELISPOT) assay in a clinical trial [41]. The study population was re-evaluated with pathology result 9 weeks after the first oral administration. Patients treated with four to six capsules per day exhibited HPV E7 specific cellular immune response in cervical lymphocytes. Seventy percent of the population showed a pathological down grade to CIN II and a correlation between disease regression and HPV E7 cellular immunity has been observed. Severe adverse effect was not reported in this study.
Listeria is a gram-positive bacteria which, unlike the gram-negative bacteria such as Salmonella does not produce endotoxin lipopolysaccharide [42]. Lm, intracellular bacterium, has two potent immunogenic components: listeriolysin O (LLO) and ActA. LLO can trigger the escape of Lm from phagosome and results in degradation of Lm by proteosome instead of lysosome. Thus, Lm can utilize both MHC class I and II pathways: The Listeria peptide in the cytoplasm processed by proteasome is displayed on MHC class I on the host cell and triggers the activation of CD8+ T cell. Alternatively, the Listeria peptide which failed to escape from the phagosome is processed and loaded onto MHC class II molecules and activates CD4+ T cell [43]. ActA can aid the formation of actin tails that assist the spreading of Lm to adjacent cells. In addition, the antigen of Lm expressed on the host cell surface can be recognized by toll-like receptor (TLR) 2 and 5, triggering the activation of innate immune responses through myeloid differentiation primary-response protein 88 pathway [44], enhancing the intensity of the subsequent antigen-specific T cell responses. Lm-LLO-E7 vaccine is a fusion of Lm, HPV16 E7 antigen, and LLO together. The phase I trial of Lm-LLO-E7 was firstly evaluated in 15 patients with recurrent, metastatic cervical cancer in clinics. Three kinds of dose were administered such as 1×109 colony-forming unit (CFU), 3.3×109 CFU, and 1×1010 CFU. Flu-like syndrome was observed in all patients, but fever and hypotension was noted only at the highest dose [45].
2) Viral-based vectors
Vaccinia virus, adenovirus, and alphavirus are examples of viral vectors that have been used in the preparation of therapeutic vaccine [464748]. Viral vectors are highly effective in entering the host cell and are very immunogenic in stimulating the T cell responses. However, viral vector has limitation for its use in immunocompromised patients due to safety concerns.
Vaccinia virus is a member of Poxviridae family with double-stranded DNA genome. It infects host cell very efficiently, controls the expression of inserted gene by specific promoter consummately, and induces the lysis of infected host cell. This characteristic reduces the chance of vaccinia virus to unexpectedly integrate into the host cell's genome [47]. The first clinical trial using live recombinant vaccinia virus, tissue antigen (TA)-HPV, expressing the E6 and E7 proteins of HPV16 and 18 was performed in eight patients with advanced stage of cervical cancer [47]. Single dose TA-HPV vaccination only generated mild and tolerable toxicity in patients. TA-HPV vaccination also induced HPV-specific cytotoxic T lymphocyte (CTL) immune response in 28% of participants (three out eight). In the trial, two patients showed tumor free condition at 15 and 21 months after TA-HPV vaccination [47].
Another TA-HPV study was performed with two times of TA-HPV vaccination in 29 patients with International Federation of Gynecology and Obstetrics stage IB or IIA who will undergo radical hysterectomy. The first dose was administered 2 weeks before surgery and the second one was injected 4 to 8 weeks after the surgery. Twenty-eight percent of patients (eight out of 29) exhibited HPV specific serological responses. However, HPV-specific CTL response was transient [49].
In another clinical study, TA-HPV was administered intramuscularly into the deltoid muscle of 12 patients age 42 to 54 years old composed of 11 patients with HPV16+ vulvar intraepithelial neoplasia (VIN) grade III and one patient with HPV16+ vaginal intraepithelial neoplasia grade II. The mean reduction of 40% in size of HPV-associated lesion was found in 83% of the population over 6 months. The significant increase of HPV16 E6 and E7-specific T cell immunity was identified via interferon γ (IFN-γ) ELISPOT assay after vaccination [50].
2. Peptide vaccine
The peptide vaccines are easy to produce, stable, safe, and little safety concern. However, poor immunogenicity of peptide-based vaccines requires the combination with immune adjuvants to enhance immune responses generated by peptide vaccine [51]. In addition, peptide-based vaccines are heavily affected by the restriction of MHC. Hence, HPV-antigenic peptides are not only selected based on the binding affinity to MHC molecule, but also its immunogenicity in eliciting CTL immune responses. For example, three immunogenic epitopes of HPV E7 were identified in human leukocyte antigen (HLA)-A2 kb transgenic mice as follow (11-20, YMLDLQPETT; 82-90, LLMGTLGIV; 86-93, TLGIVCPI) [52]. In a phase I/II trial, a total of four HPV16 E7 peptides (amino acids E7 11-20, and E7 86-93) vaccination were administered in 19 HLA-A2+ patients with cervical cancer. The vaccinations only generated mild and tolerable adverse effect, but peptide vaccinations also only elicited weak immune response [53].
Short peptides may be loaded exogenously on MHC class I of professional APC or non-professional APC through direct loading. The short peptide on MHC class I on non-professional APC can induce immune tolerance due to absence of proper costimulation. In addition, direct loading of short peptide onto MHC class I could limit MHC II presentation and prevent CD4+ T cell activation with in turn prevent the activation of CD8+ memory T cell. Compared to short peptides, long peptides cannot be exogenously displayed on MHC class I. Long peptide must be captured, internalized, and processed into small pieces before being presented on MHC class II and I. Thus, long peptide can elicit the CTL responses more efficiently than short peptide [54]. In a clinical trial, long peptide vaccines (25 to 35 amino acids) expressing HPV16 E6 and E7 were administered with Montanide ISA 51 adjuvant (Seppic Inc., Fairfield, NJ, USA) four times in 35 patients with advanced cervical cancer [55]. IFN-γ+ T cell responses after coinjection of overlapping long peptide were predominantly specific against HPV E6 peptide. Severe adverse toxicity was not observed.
To overcome the low immunogenicity of peptide vaccine, 4–1BB ligand [51] and bryostatin-I [56] have been adopted as costimulatory material. Costimulatory molecule is indispensible for the activation of innate and adaptive immune system. 4–1BB ligand can enhance the uptake of DC and stimulated more cytotoxic and memory T cell [51]. Bryostatin-I can induce chemokine (C-C motif) ligand 2 release from DC, which plays a very important role in trafficking of DC and lymphocyte into lymphoid organ [56]. Poly (I:C), a synthetic RNA that can activate DC via TLR 3 [57], and monoclonal anti-CD40 antibody that can stimulate interaction between DC and CD8 T cell [58] were incorporated in TriVax peptide vaccination containing HPV16 E749-57 [59].
3. Protein vaccine
Protein vaccine can overcome the MHC restrictions that represent a significant concern for peptide vaccine. All possible epitopes can be derived from protein vaccine via processing by APCs. Hence, protein vaccine dominantly leads to more antibody-production than cytotoxic T cell activation. Similar to peptide vaccine, protein vaccine also requires immune stimulatory adjuvant to surmount low immunogenicity [40].
SGN-00101, whole sequence of HPV16 E7 combined with heat shock protein (HSP) 65, was subcutaneously administered three times to 58 patients with CIN III in a clinical trial [60]. Clinical responses were determined on the surgical specimen after loop electrosurgical excision procedure or cone biopsy. Seventy-eight percent of the population showed clinical responses (complete response, partial response), while two patients (3.5%) progressed to micro invasive cervical cancer. No significant drug related toxicity was reported.
Tissue antigen-cervical intraepithelial neoplasia (TA-CIN) is an 80 kD L2E7E6 monomer, 725 amino acids long protein based vaccine [61]. In preclinical study, TA-CIN vaccination in tissue culture-1 (TC-1) tumor bearing mice generated HPV16-specific antibodies and CTL responses, translating into the protective and therapeutic potential of the vaccine. Phase I trial was performed in 40 healthy volunteers to evaluate safety and immunogenicity of TA-CIN [62]. TA-CIN was administered intramuscularly three times in 4-week interval. Importantly, HPV16 E6 and E7 specific responses were detected through ELISPOT assay with little to no adverse events observed.
Imiquimod is the first immune modulator and vanishing cream with oil-in-water structure. It has been widely used to treat genital warts [63]. It acts as agonist for TLR 7 and 8 and activates nuclear factor κB (NF-κB), which induces Th1-dominant immune response. It has also been shown to induce the trafficking of Langerhans cells to draining lymph nodes and the maturation of DCs [64], as well as inhibiting myeloid-derived suppressor cells (MDSC) in the tumor microenvironment that have immunosuppressive functions [63]. Phase II trial with imiquimod and TA-CIN was conducted on 19 patients with VIN II or III [65]. Imiquimod was applied on VIN lesion for 8 weeks and then TA-CIN was administered three times in 4-week interval. Complete remission of VIN was observed 63% of the patients (12 out of 19) and significantly more CD4+ and CD8+ T lymphocytes infiltration into the lesion were observed in responders compared to non-responders.
4. DNA vaccines
The advantages of DNA vaccine include safety, ease of production, stability, and ability of repeated vaccination without inducing neutralizing antibody. Target gene inserted in plasmid DNA is transfected into host cell and the final protein are produced through transcription and translation. It can stimulate cellular and humoral immunity through MHC class I and II. However, DNA vaccines generate relative low immunogenicity and are necessary to be boosted up with the help of adjuvant methods such as delivery tool or immune stimulator [33].
To achieve high efficacy for DNA vaccine, the following conditions are required. Firstly, sufficient amount of DNA material need to be introduced via intramuscular or intradermal route to stimulate immune cell. Secondly, sufficient numbers of APC are present to uptake the target antigen. Thirdly, efficient antigen processing and presentation by APC is necessary. Lastly, full activation of T cell is mediated by APCs [4066].
After intramuscular injection of DNA vaccine, the DNA may be taken up by either myocytes of DCs. Myocytes that take up the DNA will express and secret the encoded antigen, which will then be picked up by DC through phagocytosis and expressed on MHC I complex through cross presentation [67]. Alternatively, the DCs directly transfected by vaccination will express, process, and present the antigen to CD8+ T cell in the draining lymph node via direct presentation pathway [68].
In one of the clinical trials evaluating the efficacy of therapeutic HPV DNA vaccine, the DNA vaccine, pNGVL4a-CRT-E7 (detox), was administered three times to 32 patients with CIN II and III patients [69]. Three delivery methods were tested in the study: particle mediated epidermal delivery, intramuscular injection, and intralesional administration. Thirty percent of patients (eight of 27) who received all vaccinations showed clinical regression to CIN I. Intralesional administration induced significant infiltration of CD8+ T cell into cervical dysplasia. Twenty-two of the 32 enrolled patients experienced vaccine-related adverse events, but all observed adverse events were grade 1 or less in severity and were well tolerated.
In DNA vaccine pNGVL4a-CRT-E7 (detox), the sequence of HPV16 E7 was combined with calreticulin (CRT) to enhance immune response of CD8+ T cells. CRT is a members of HSP, a category of proteins that are expressed in the condition of cell stress such as heat or starvation and composed of HSP70, HSP90, HSP110, and CRT, and facilitates the transportation of MHC I-peptide complexes into endoplasmic reticulum. In addition, CRT also showed anti-tumor effect through selective inhibition of proliferation of vascular endothelial cell [70].
In another clinical trial, GX-188, a therapeutic HPV E6 and E7 DNA vaccine, was administered in nine patients with CIN III [71]. In this trial, electroporation was combined with DNA vaccine to increase the immunogenicity of the therapy. In addition, Fms-like tyrosine kinase-3 ligand was inserted into plasmid vector to facilitate antigen presentation on DCs. The vaccination stimulates the differentiation of CD4+ T cells into Th1 cells. Importantly, eight out of nine patients exhibit enhanced multifunctional HPV16-specific CD8+ T cell immune responses and 77% of the population (seven out of nine) showed complete regression of CIN III lesions as well as viral clearance after the completion of treatment.
Electroporation technique utilized in the GX-188 trial is a DNA vaccine delivery method that can increase the transfection efficiency of the vaccination. The electroporation induces an electric current that transiently augments the cell membrane and aid DNA uptake into cell. In addition, electroporation generate local inflammation which facilitates the recruitment APCs to vaccination site [72].
Electroporation has been utilized for the administration of multiple therapeutic DNA vaccines. A separate clinical trial evaluated the efficacy of intramuscular injection of VGX-3100 (Inovio Pharmaceuticals, Plymouth Meeting, PA, USA), a therapeutic HPV DNA vaccine encoding HPV16 and HPV18 E6/E7, combined with electroporation in eighteen women previously treated for CIN II or III [73]. No serious adverse events associated with vaccination have been reported. Moreover, 56% of the patients (10 out of 18) generated serum antibodies specific for all types of VGX-3100 antigens. Also, the induction of functional CD4+ and CD8+ T cells responses were observed after vaccination. Besides electroporation, other delivery methods to enhance the efficacy of DNA vaccines have been developed, including gene gun, laser treatment, and microencapsulation which exploited the aggregation of APC into the application area [40].
5. Dendritic cell-based vaccines
DCs, which are the most efficient professional APCs, have important regulatory roles in the immune system. DCs activate innate and adaptive immunity against HPV infection by generating diverse proinflammatory cytokines. They efficiently process and present antigens to T cells through both the MHC class I and MHC class II pathways. DC-based HPV vaccines using HPV16 and/or HPV18 E7 have been shown to induce significant E7-specific CD4+ and CD8+ T cells immune responses in patients with cervical cancer [74]. Full-length E7-pulsed autologous DCs induced E7-specific CD4+/CD8+ CTL responses and lysed HPV-infected cancer cells in cervical cancer patients [75].
In addition to introducing antigens into DCs, strategies can be incorporated into DC vaccines to increase the efficiency of the vaccine. One such example includes the incorporation of shRNA against suppressor of cytokine signaling 1 (SOCS1) [76]. SOCS1 inhibits the Janus kinase/signal transducers and activators of transcription signaling pathway as part of a negative feedback loop and can suppresses the function of IFN-γ, inhibits Th1 responses, DC activation, and negatively regulates the TLR–NF-κB pathway [77]. A study used adenovirus to introduce shRNA-SOCS1 into HPV16 E7-pulsed DC vaccine, and showed that silencing SOCS1 in antigen-presenting DCs can positively facilitate antigen-specific antitumor immunity, resulting in generation of more potent anti-tumor effect in TC-1 tumor-bearing mice [76].
INHIBITOR OF IMMUNE CHECKPOINT IN THE IMMUNOTHERAPY OF CERVICAL CANCER
For the balance of immune system, cellular immunity is regulated by both activation signals (costimulatory molecules) as well as inhibition signals (immune checkpoints) [78]. In normal condition, immune checkpoints are responsible for maintaining self-tolerance, preventing autoimmunity, and protecting normal cells from immune attack during infections. However, cancer cells often exploit this mechanism to their advantage to escape from immune surveillance. Indeed, immune suppressive milieu is observed in abundance inside tumor microenvironment. Hence, inhibition of immune checkpoint is suggested as one potential method to enhance anti-tumor immunity.
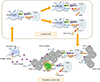
Among a variety of immune checkpoint, cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) and programmed death receptor (PD-1) were few powerful immune suppressor being intensively studied [7980]. CTLA-4 begins to be expressed on the naïve T cell 48 hours after activation in lymph node and is closely associated with attenuation of these activating T cells [81]. On the other hand, PD-1 is expressed on effector T cell in peripheral tissues and binds with programmed death ligand 1 (PD-L1, B7-H1) and PD-L2 (B7-DC) expressed on DCs or tumor cells for attenuation of activated effector T cell (Fig. 3) [8283]. In normal condition, PD-L1 is induced by IFN-γ and serves to protect DCs from T cell mediated cytotoxicity [80]. In HPV associated head and neck cancer, a higher percentage of PD-1 expressing CD8+ T cell was observed in tumor tissue compared to in the peripheral blood, it implies that CD8+ T cells begin to express PD-1 receptor after entering tumor environment.
Fig. 3
The function of immune checkpoint inhibitor in tumor. Tumor antigen is captured by dendritic cell and delivered to naïve T cell in lymph node. Normally, immune checkpoint (cytotoxic T-lymphocyte–associated antigen 4 [CTLA-4]) on activated T cell is expressed to keep the balance of immune system. Anti-CTLA-4 antibody blocks the inhibitory signal of T cell and extends the duration of activated T cell. Programmed death ligand 1 (PD-L1) expressed on periphery tumor site prohibits activated T cell through programmed death receptor (PD-1) receptor expressed on effector T cell. Anti-PD-1 antibody eliminates the interaction of PD-1 and PD-L1 and aids to keep effector T cell activated enough to kill tumor cells. MHC, major histocompatibility complex; TCR, T cell receptor.
Human antibody against CTLA-4, Ipilimumab (Bristol-Myers Squibb, Princeton, NJ, USA), was approved by U.S. Food and Drug Administration (FDA) in 2011. Ipilimumab administration has been shown to improved survival of metastatic melanoma patients [81]. Absolute lymphocyte count (ALC) and CD8+ T cell are suggested to monitor the results after Ipilimumab treatment. ALC >1,000/µL after two times Ipilimumab was significantly related with clinical remission and increase of overall survival [84]. In 2014, FDA approved humanized antibody against PD-1 (Pembrolizumab, Merck & Co. Inc., Whitehouse Station, NJ, USA) as well as human anti-PD-1 (Nivolumab, Bristol-Myers Squibb) for the treatment of advanced melanoma [8586]. Multiple clinical trials have been planned and conducted to incorporate these therapeutic agents into treatment regimens against HPV-associated cancers.
A phase I trial of Ipilimumab following CCRT (NCT01711515) is currently undergoing in patients with locally advanced cervical cancer. Ipilimumab will be intravenously administered to patients every 3 weeks for 3 months. Also, a phase II trial is currently testing the efficacy of Nivolumab in treating patients with persistent, recurrent, or metastatic cervical cancer (NCT02257528).
Other cells and molecules in the tumor microenvironment other than check point inhibitors that have immune inhibiting functions includes regulatory T cell [87], MDSC [88], indoleamine 2, 3-dioxygenase [89], arginase [90], nitric oxide synthase [91], IL-4–induced gene 1 [92], IL-10 [93], and TGF-β [8894]. The ability to overcome these immune suppression may further enhance the potency of immunotherapy against HPV-associated cancers.
CONCLUSIONS
In conclusion, a number of HPV therapeutic vaccines have been developed to enhance the function of DC and T lymphocyte for the treatment of cervical cancer and HPV-associated diseased. Recently, immune checkpoint inhibitors were incorporated into treatment modality and clinical trial. These therapeutic agents have the potential to drive down disease burden associated with HPV-infection, and improve the living quality of the patients.
 XML Download
XML Download