

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
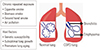
Chronic obstructive pulmonary disease (COPD) is characterized by persistent respiratory symptoms and airflow limitation due to airway and/or alveolar abnormalities usually caused by prolonged exposure to noxious particles or gases.1 Cigarette smoking is the most common cause of COPD; however, environmental factors, such as polluted air, second-hand smoke, or biomass smoke, could also cause COPD. In addition, genetic factors or suboptimal lung growth in childhood may be associated with the development of COPD (Fig. 1).2 Currently, COPD is the fourth leading cause of deaths worldwide, and by 2020, it is expected to be the third leading cause of deaths worldwide.1 Approximately three million people died of COPD in 2015, accounting for 5% of all deaths worldwide in that year.3 Due to the high prevalence and chronicity of COPD, it has been a global burden for many years, and the mechanisms of its development remain unidentified. Intriguingly, the incidence of COPD becomes higher with aging, and peak incidence is observed in patients aged 65–74 years.45 Therefore, aging is considered a critical factor in the development of COPD.6 It has been proposed that accelerated premature aging of the lungs may be the mechanism of COPD development.7 Once a patient is suspected as having COPD, the diagnosis is confirmed based on spirometry results. For diagnosis of COPD, patients should have irreversible airflow limitation, which is defined by post-bronchodilator forced expired volume in 1 second (FEV1)/forced vital capacity (FVC) ratio of <0.70 according to the Global Initiative for Chronic Obstructive Lung Disease (GOLD) guidelines.8 It is well documented that lung function typically declines with aging. Specifically, FEV1 decreases, and aging lungs, just like COPD-affected lungs, often have FEV1/FVC ratio <0.70 and increased residual volume.9 This is because the physiology of aging lungs and COPD-affected lungs are similar, showing decreased elastic recoil of the lungs, stiffened chest wall, and altered gas exchange.6 In addition, similarly to COPD-affected lungs, aging lungs have decreased mucociliary clearance, compromised vascular regulation, and alveolar enlargement.10 However, alveolar enlargement in aging lungs, so-called “senile emphysema,” differs from that in COPD in terms of the lack of alveolar wall destruction and inflammation.9 It has been suggested that the main driving force for COPD development could differ among individuals. For instance, suboptimal lung growth in childhood has also been shown to play an important role in causing COPD in some individuals.11 Taken together, although premature lung aging may not be a pathogenic mechanism in all COPD patients, many aspects of aging lungs and COPD-affected lungs are similar, implying that the mechanisms of aging may contribute to the development of COPD. In this study, based on our literature review, we propose that immunosenescence, which is the gradual deterioration of the immune system brought on by natural age advancement, is the key pathogenetic mechanism of COPD.
COPD AS A CHRONIC INFLAMMATORY DISORDER
COPD is a chronic inflammatory disease induced by repeated exposure to respiratory irritants, mostly cigarette smoke, and inflammation in COPD-affected lungs continues to be observed even after stopping smoking.12 Normal immune responses during inflammatory processes are expected to prevent lungs from further injury or infection; however, chronic inflammation in COPD-affected lungs is associated with abnormal immune responses leading to increased tissue damage, resulting in remodeling of the lung.12 Inflammation in COPD is predominantly driven by type 1 immunity and type 3 immunity.13 Both innate and adaptive immune systems are involved in the development of chronic inflammation leading to COPD. The role of innate immunity in COPD pathogenesis has been extensively studied recently in terms of the induction and acute exacerbation of the disease.1114 Interleukin (IL) 1-like cytokines are reportedly elevated in the lungs of COPD patients, implying the role of an inflammasome in the pathogenesis of COPD.1114 The inflammasome is a multiprotein intracellular complex that activates proinflammatory caspases, such as caspase 1, leading to the cleavage of proinflammatory cytokines and production of active IL-1β and IL-18 in response to pathogenic microorganisms and sterile stressors.1114 To activate the inflammasome, innate immune cells should recognize pathogen- associated molecular patterns (PAMPs) of microbial pathogens or damage-associated molecular patterns (DAMPs) of sterile stressors that are categorized based on their subcellular origin (e.g., cytoplasm, endoplasmic reticulum, nucleus, and mitochondria). This recognition of PAMP or DAMP is achieved through several families of pattern recognition receptors (PRRs) expressed in innate immune cells.1114 The PRRs include Toll-like receptors (TLRs), nucleotide-binding domain leucine-rich repeat-containing receptors (NLRs), C-type lectin receptors, and RIG-I-like receptors.1114 In the lungs, TLRs are expressed on various cells, including alveolar macrophages, lymphocytes, dendritic cells, and bronchial epithelial cells.1114 Among different TLRs, TLR4 reportedly plays an important role in murine models of COPD.151617 TLRs recognize molecular patterns on the cell surface of a microbial pathogen, whereas NLRs identify molecular patterns in the cytosol of the host cell.18 It has been shown that the DAMPs generated by cigarette smoke-induced death of airway epithelial cells or the PAMPs induced by microbial antigens activate the NLR family pyrin domain containing 3 (NLRP3) inflammasome, leading to the release of IL-1β with subsequent neutrophilic inflammation in the lungs of COPD patients.1920 However, the role of the inflammasome in COPD development has been challenged recently. One study reported that, although innate immunity reportedly correlates with disease progression in COPD patients, the NLRP3 inflammasome is not activated in patients with stable COPD,21 thus, the exact roles of innate immunity, including that of the inflammasome, in COPD pathogenesis remain unclear. Several studies have reported on adaptive immune responses in the lungs of COPD patients. Active participation of cytotoxic CD8+ T cells, T helper 1, and 17 CD4+ cells and B-cell response leading to the production of antibodies have been reported in COPD-affected lungs.22 Upon activation, CD8+ T cells release proteolytic enzymes, such as perforin or granzymes, which cause cell death of structural cells by apoptosis or necrosis.22 Th1 cells secrete interferon gamma and Th17 releases IL-17, thus leading to increased inflammation at the site of injury in response to cigarette exposure.2324 In addition, smokers with COPD have significantly fewer T-regulatory cells in the lungs; this may contribute to the perpetuation of inflammation and the induction of autoimmunity. The development of autoimmunity has been mostly reported in patients with severe COPD.25 Further, B cells are also increased in large airways, and peribronchial lymphoid follicles are often observed, which are due to lymphoid neogenesis in COPD-affected lungs.26 Taken together, it is very likely that the interaction between innate and adaptive immune cells may orchestrate the onset, progression, and severity of lower airway inflammation in COPD.21
IMMUNOSENESCENCE
Given that COPD, a chronic inflammatory lung disease, is probably caused by accelerated lung aging, the changes in immune systems with aging may be associated with COPD pathogenesis. The immune system also undergoes an aging process termed immunosenescence. Both innate immunity and adaptive immunity are affected by aging.27 Aging innate immune cells, such as neutrophils, macrophages, dendritic cells, and natural killer cells, undergo functional decline in phagocytosis, chemotaxis, their ability to secrete inflammatory cytokine, antigen- presenting capacity, and bactericidal ability.28293031 Declining function of innate immune cells with aging also contributes to the dysregulation of the adaptive immune system via molecular cross-talk.32 The decline in cell-mediated immunity forms a part of the aging immune profile, which is primarily characterized by reduced thymic output leading to reduced T-cell repertoire and increased oligoclonal expansion of memory cells.33 In other words, reduced responsiveness to new antigen load, owing to the reduced naïve to memory cell ratio and expansion of mature cell clones, characterizes immunosenescence of cell-mediated immunity.34 Further, aging is also associated with shifts from Th1 cytokine profile to Th2 cytokine profile in response to immune stimulation. The overproduction of Th2 cytokines in this setting may augment B-cell-mediated autoimmune disorders in older adults.35 Humoral immune function also changes significantly with aging; these changes include decreased antibody responses and diminished production of high-affinity antibodies related to defective surface immunoglobulin/B-cell receptor affinity, decreased signaling, and reduced B-cell proliferation.36 There is also a loss of naïve B-cells and an increase in memory cells with age,37 resulting in a reduced ability to respond to new antigens.38 Overall, the immunosenescence of adaptive immunity results from the depletion of the reservoir of naïve cells over time as a result of repeated exposure to pathogens and their conversion to memory cells.33
INFLAMMAGING (CHRONIC INFLAMMATION)
Immunosenescence leads not only to impaired immune responses but also to low-grade, chronic, systemic inflammation, which is described as inflammaging.39 Inflammaging is characterized by elevated proinflammatory cytokines, such as IL-1, IL-6, and tumor necrosis factor (TNF)-α.40 Inflammaging is believed to be a cumulative result of lifetime exposure to antigenic load from infections, as well as exposure to noninfectious antigens.41 Key genes that are known to regulate inflammaging include the insulin-like growth factor (IGF-1) signaling pathway, target of rapamycin (TOR), and AMP-activated protein kinase (AMPK).42 Not surprisingly, inflammaging has been proposed as one of the contributors to the development of most age-related disorders and as a common biological factor responsible for the decline and onset of diseases in older adults.4344 Many factors are likely involved in inflammaging. First, intrinsic dysregulation of immune cells simply may be a cause of inflammaging. Regarding the innate immune system, monocytes and macrophages are suggested to contribute to inflammaging significantly by the active generation of inflammatory cytokines.29 In addition, T cell-mediated immunity can induce inflammaging by changing their phenotype into a more proinflammatory phenotype in a manner dependent on previous exposure, reactivation of antigenic challenges, and exhaustion of T-cell repertoire.29 The CD8+ T cell population is thought to be a bigger contributor in this regard than the CD4+ population.45 Second, senescent cells can produce proinflammatory mediators and proteases, which are collectively termed as senescence-associated secretory phenotypes (SASPs).46 SASPs may activate inflammatory responses by recruiting immune cells possibly to cause inflammaging.47 Third, age-related changes in adiposity and hormones (e.g., estrogen) reportedly cause inflammation. Proinflammatory cytokines and free fatty acids secreted from inflamed adipose tissue can activate inflammation, which may be a part of inflammaging.4849 Fourth, age-associated changes in gut microbe concentrations may foster an imbalance in vivo that affects inflammaging.50 Fifth, increased cell death or damage to cellular components induced by cellular senescence and age-related stress, such as metabolic stress, may exceed the capacity limit of the phagocytic system of aged myeloid cells, resulting in the accumulation of DAPMs and the activation of inflammasome.5152 This process can be further intensified as aging is associated with decreases in autophagy.53 Autophagy is a cellular housekeeping mechanism responsible for the removal of dysfunctional and damaged intracellular proteins via lysosomal degradation. Thus, one of the roles of autophagy is to prevent the activation of inflammasomes. The consequence of a decline in autophagy with aging is therefore increased activation of the inflammasome and greater proinflammatory responses.54 All these abovementioned age-related changes can contribute to inflammaging among older adults.
COPD AND INFLAMMAGING
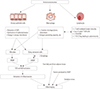
Given that inflammaging in the context of immunosenescence has been implicated in the pathogenesis of most age-related chronic diseases, COPD could be one of the target diseases of inflammaging.4344 Previous study demonstrated that aged mice developed more inflammation leading to emphysema when exposed to cigarette smoke, compared to younger mice.55 If inflammaging plays a significant role in the generation of COPD, the specific mechanisms for inflammaging-induced COPD need to be explored. Airway epithelial cells seem to be a key player in this regard. Airway epithelial cells are pivotal innate immune cells and serve as a physical and molecular barrier for particulate matter against microbes, as these cells entrap all dangerous molecules via mucociliary clearance, and then, these molecules ultimately get degraded in the gastrointestinal tract. By this process, airway epithelial cells can prevent the access of molecules that can activate TLR deployed on their basolateral surface. Once TLR agonists breach this epithelial barrier, they become critical danger signals for activating the strategically arrayed cellular components of the innate immune defense.56 The airway epithelial cells can serve as a physical and molecular barrier as they are polarized and maintain a tight junction that prevents paracellular transport of microbes and harmful substances into underlying lung tissues in normal conditions. Aging is associated with decreased epithelial barrier function,57 abnormalities in both cilia structure and function,58 and reduced production of antimicrobial and anti-inflammatory peptides produced by epithelial cells, including secretory leukocyte protease inhibitor (SLPI).59 Not surprisingly, both alveolar and airway epithelial cells of smokers with COPD reportedly have increased numbers of senescent cells relative to healthy controls.60 Another key cell in terms of the generation of inflammaging in the lungs might be the airway and alveolar macrophages. Communication between airway epithelial cells and airway macrophages seems to be critical for immune homeostasis within the lungs. For instance, airway epithelial cells and airway macrophage cooperate to phagocytose particulates or microorganisms.61 Aging macrophages undergo the functional decline in phagocytosis, chemotaxis, ability to secrete inflammatory cytokine, antigen-presenting capacity, and bactericidal ability.28293031 Therefore, the capability to protect the lungs against inhaled particles or infectious agents will be decreased with aging, resulting in more inflammation in the lungs. Recently, changes in the airway microbiome have been implicated in various lung diseases.62636465 Due to difficulties in sampling the airways while preventing contamination by concurrent oropharyngeal microbes, understanding of the airway microbiome was initially not easy; however, a series of studies utilizing DNA sequencing approaches have demonstrated the microbiota of the lower airways. Further, it is now clearly accepted that variations in bacterial abundance, content, and structure occur in chronic inflammatory airway diseases, such as cystic fibrosis,62 COPD,63 and asthma.6465 This means that a particular microbiome may be characteristic of certain airway pathology. Taken together, the interaction between the microbiome and the host immune system within the airway, as well as the consequent loss of immune function with age, may have implications for the onset and progression of various age-related chronic airway diseases.66 Additionally, increased oxidative stress has been strongly implicated in both COPD and aging lungs as determined by reactive oxygen species levels, oxidized DNA, lipid peroxidation and nitric oxide levels.6768 Certainly, this increased level of oxidative stress can contribute to the development of inflammaging. Other factors associated with inflammaging, which have been described earlier, including intrinsically abnormal senescent immune cells, accumulation of DAMPs from the dying senescent cells in the setting of decreased autophagy in aging cells, or SASP produced by senescent cells, can also contribute to the development of inflammaging in the lungs of smokers, thus, possibly, leading to COPD. As described above, aging lungs seem to be prone to developing inflammation in response to certain stimuli irrespective of whether they are related to inflammaging or not. From the COPD standpoint, chronic inflammation can develop when inflammation-prone aging lungs are repeatedly exposed to respiratory irritants, such as cigarette smoke, ultimately leading to tissue destruction and the development of COPD. Notably, COPD patients often undergo acute exacerbation. In the cases of acute exacerbation of COPD, which is typically caused by a viral or bacterial infection, chronic inflammation can be further intensified.69 Intriguingly, only 15% to 20% of smokers develop clinically significant COPD. Thus, the majority of smokers do not develop COPD.70 This is different from the mouse model of emphysema, which gives a predictable dose-response relationship between smoking amount and emphysema.70 Currently, we do not know the reason behind only a small percentage of smokers developing COPD. If immunosenescence and/or inflammaging play a critical role in the development of COPD, the reasoning for COPD occurring in a small percentage of smokers can perhaps be explained from immunosenescence or inflammaging or both perspectives. There is reportedly a huge inter-individual variation in inflammaging susceptibility, which seems to be, in part, genetically determined through polymorphisms of promoter regions of cytokine genes or the length of telomeres.29 A negative correlation between telomere length and the levels of C-reactive protein71 and IL-672 has also been observed, and shorter telomere length in leukocytes from COPD patients has been demonstrated.73 In addition, enhanced telomere erosion has been observed in the small airway epithelium from COPD patients.74 Therefore, it is quite possible that susceptibility to inflammaging, which is genetically determined, may explain the inter-individual vulnerability to the development of COPD among smokers. In addition, thymic involution, a factor associated with immunosenescence, begins early in life, and the size of the naïve cell pool is mostly determined before puberty.33 Thus, it is possible that immunosenescence is significantly affected by the history of an individual's exposure to pathogens. In other words, the cumulative effects of antigenic stressors may determine immunosenescence or inflammaging or both and thus explain the differences among smokers in their susceptibility to the development of COPD (Fig. 2).
CONCLUSION
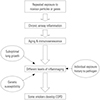
Based on our literature review, we have proposed immunosenescence and inflammaging as the driving mechanisms of the development of COPD and have outlined why only a minority of smokers develop COPD (Fig. 3). If this is true, we could slow the progression of COPD by regulating immunosenescence and inflammaging in the lungs. Future studies will be needed to support our hypotheses.
 XML Download
XML Download