

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
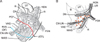
The rectovaginal septum covers the posterior aspect of the vagina, whereas the pubocervical fascia covers the anterior aspect; both fasciae provide so-called level II support to the female pelvic viscera, preventing prolapse [1]. Mostwin [2] considered the pubocervical fascia to be a tissue filling the space between the vagina and the urethra or bladder. The tendinous arch of the pelvic fascia (arcus tendineus fasciae pelvis) has been a common target in studies of gross anatomy [3, 4, 5]. Along the tendinous arch, the rectovaginal septum and pubocervical fascia meet the superior fascia of the levator ani, the two structures being apparently embedded in the lower paracolpium or the paravaginal tissue [1]. The rectovaginal septum corresponds to the Denonvilliers' fascia in women [6]. Because the mesorectum (a fatty tissue layer surrounding the lower rectum) is adjacent to the Denonvilliers' fascia in women [7], the covering fascia of the mesorectum most likely joins, or correspond to, the rectovaginal septum. These features of the fasciae along and around the vagina are summarized in Fig. 1.
Donker [8] appers to have been the first researcher to have noted abundant thick nerves running through the paracolpium from the pelvic plexus to the urologic organs, and his observations were followed up by Ball et al. [9]. Thus, the paracolpium is likely to include at least some of the pelvic autonomic nerve plexus (Fig. 1). Using late-stage fetuses, Kato et al. [10] demonstrated the female cavernous nerve running through the putative paracolpium. Hirata et al. [11] provided an elegant diagram showing the course of the female cavernous nerve covering the pubocervical fascia, but they did not provide a photographic figure of the nerve. To identify the cavernous nerve, immunohistochemistry for neuronal nitric oxide synthase (nNOS) is necessary [12]. Using sections from a 21-week-old fetus, Alsaid et al. [13] demonstrated and reconstructed the female cavernous nerve running alongside the rectum and vagina. However, reports describing the application of nNOS immunohistochemistry to the adult pelvic nerves seem to be very limited, possibly because of difficulty in obtaining good materials for the immunoreaction [14]. Likewise, the application of immunohistochemistry for vasoactive intestinal peptide (VIP) in adult tissues seems to have been limited to a study of intramural nerves in the vagina [15].
Any dissection study designed to observe the topographical anatomy including nerves and fasciae in the pelvis may be limited because of the delicate architecture: minute dissection of autonomic nerves [16, 17] may not be appropriate for demonstration of pelvic fasciae, as was shown in two elegant studies of fresh cadavers published in the same year [18, 19]. There have been several attempts to show both the nerves and the fascia: using frozen cadavers, Hollabaugh et al. [20] demonstrated an "intrapelvic" course of the female pelvic splanchnic nerves alongside the extrapelvic pudendal nerve as a result of broken fasciae. Likewise, Mostwin [2] demonstrated these nerves through windows artificially created in the fasciae. Conversely, histological descriptions of female topographical anatomy among the fasciae and intrapelvic nerves along the tendineous arch require large sections that include related structures, and this seems to have limited the number of such studies [6, 21]. Although large sections are good for demonstration of the pelvic fasciae, but they are not suitable for immunohistochemistry because of their thickness and/or acidic decalcification. Therefore, using classical paraffin sections, the aim of the present study was to demonstrate in detail the topographical anatomy of both the nerves and fasciae in the paracolpium with the aid of immunohistochemistry.
Materials and Methods
This study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We examined 10 donated female cadavers ranging in age from 73 to 100 years, with a mean age of 85 years. The cadavers included two nulliparous women (Table 1). In all cases the cause of death had been ischemic heart failure or intracranial bleeding and we confirmed that none of the women had undergone surgery by reference to their individual histories as well as macroscopic observation after opening the abdominopelvic cavity. These cadavers had been donated to Sapporo Medical University for research and education on human anatomy, and their use for research had been approved by the university ethics committee. The donated cadavers had been fixed by arterial perfusion of 10% v/v formalin solution and stored in 50% v/v ethanol solution for more than 3 months. From each cadaver, we prepared one large tissue block including the bladder, urethra, uterus, vagina, rectum, levator ani muscle, and any connective tissue around these viscera. Five to seven slices (mostly 15 mm in thickness) were prepared from each of the hemiblocks after bisection along the midsagittal line, and after performing routine procedures for paraffin-embedded histology, five to seven large sections (70×50 mm) stained with hematoxylin and eosin (H&E) were prepared at 2-3-mm intervals from each of the slices. The sectional planes corresponded to tilted frontal planes according to Tamakawa et al. [21] in order to show the bladder, uterus (or vagina), and rectum together in a single plane. After observation of the large sections, we prepared normal-size sections for immunohistochemistry near the former large section. Thus, from one paraffin block containing a 15-mm-thick slice, we prepared 3-4 large sections and 15-20 normal-size sections.
Most sections were stained with H&E and some were used for elastica Masson staining for elastic fibers [22, 23] and immunohistochemistry. The primary antibodies used for nerve immunohistochemistry were 1) mouse monoclonal anti-human S100 protein (1:100, Dako Z0311, Dako, Glostrup, Denmark), 2) rabbit polyclonal anti-human neuronal nitric oxide synthase or nNOS (1:100, Cell Signaling Technology, Beverly, MA, USA), 3) mouse monoclonal anti-superolateral part, while nNOS-andhuman vasoactive intestinal peptide (H-6) or VIP (1:100, sc25347, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and, 4) rabbit polyclonal anti-human tyrosine hydroxylase (TH; 1:100, ab152, Millipore-Chemicon, Temecula, CA, USA). Rather than PGP9.5, neuron-specific enolase and PMP22, we have found that S100 protein is the best marker in long-preserved cadaveric specimens [10, 24, 25, 26]. In addition to elastica Masson staining, to understand the connective tissue architecture, we used mouse monoclonal anti-human alpha smooth muscle actin (1:100, Dako M0851, Dako) as the primary antibody. For all primary antibodies, the secondary antibody was labeled with horseradish peroxidase (HRP), and antigen-antibody reactions were detected by the HRP-catalyzed reaction with diaminobenzidine. Counterstaining with hematoxylin was performed on the same samples. A negative control without a first antibody was set up for each of the specimens. Observations and photography were usually performed with a Nikon Eclipse 80 (Nikon, Tokyo, Japan), but photos at ultralow magnification (less than ×1 at the objective lens) were taken using a high-grade flat scanner with translucent illumination (Epson scanner GTX970, Tokyo, Japan).
Results
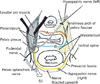
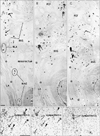
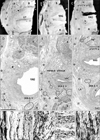
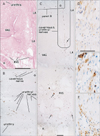
The present large sections covered a wide area including the anal sphincter at the posterior end, the urethra at the anterior end, the ureter at the superior end, and the levator ani muscle and ischiorectal fossa along the lateral margin. The retrorectal course of the hypogastric nerve as well as the proximal course of the pelvic splanchnic nerve lay outside the sections. Lower-magnification histology, shown in Figs. 2, 3, 4, 5, 6, corresponded to areas almost 2/3 of the sectional area. Figs. 2 and 3 (or 4, 5, 6) demonstrate the morphology of nuliiparous (or multiparous) women. For convenience, macroscopic slices before histological procedures are shown at the top of Figs. 2 and 4, although the histological sections display planes deeper than the surface of the slice. Using elastica-Masson staining as well as immunohistochemistry for alpha smooth muscle actin, components of the connective tissue fibers are shown at the bottom of the same figures. Likewise, for nerve fibers in the paracolpium (Figs. 3, 5, 6), immunoreactivity for nNOS and VIP was used for indicating parasympathetic nerves and that for TH was used for indicating sympathetic nerves. We used Auerbach's plexus of the rectum for monitoring any false-negative reaction for nNOS or VIP (figures not shown). As a result, nNOS (or VIP) reactivity in the rectum was identified in 7 (or 8) of the 10 cadavers, but not in the other 3, possibly due to poor preservation after death (Table 1). In contrast, reactivity for S100 or TH was consistently observed.
The rectovaginal septum (Denonvilliers' fascia) was consistently seen as a fibrous boundary facing the mesorectal loose tissue in the medial area near the midsagittal line, but it was fragmented or unclear in the lateral area (i.e., the paracolpium) in 5 specimens (multiparous or uniparous women) (Fig. 4E). The rectovaginal septum was composed of a mixture of collagenous and elastic fibers (Fig. 4I). The pubocervical fascia was consistently unclear in the medial area, but identifiable as a rough fibrous network of thick fiber bundles in the lateral area. The latter fascia provided a boundary for the paracolpium facing the thick subperitoneal fatty tissue (Fig. 2D-F). The pubocervical fascia and the rectovaginal septum did not contain smooth muscles (Fig. 2G, H). In contrast, the superior fascia of the levator contained or accompanied smooth muscles at sites near the rectum (Fig. 2I), the smooth muscles being continuous with the adjoining longitudinal muscle coat of the anal canal.
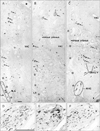
Notably, in 3 of the 10 cadavers examined, we found definite fascial connections at the tendineous arch of the pelvic fasciae; therein, the rectovaginal septum and the pubocervical fascia were each independently connected with the superior fascia of the levator ani (nulliparous, 2; multiparous, 1) (Fig. 2E, F). In these 3 cadavers, nerves in the paracolpium were almost evenly distributed to form a sheet-like configuration; at levels below the uterine cervix, the density was higher in the lateral area near the superior fascia of the levator ani than in the medial area (Fig. 3A-C). The nerves were embedded in elastic fiber-rich tissues. In a single cadaver (uniparous), any clear fascial connection was restricted to the upper part of the vagina. In another cadaver (multiparous) (Fig. 6A), the rectovaginal septum was connected with the superior fascia of the levator ani, but the pubocervical fascia appeared to be replaced by the venous plexus. The venous plexus was embedded in elastic fiber-rich, loose tissues of the paracolpium (Fig. 4H). In the other 5 cadavers, the fascial connection at the tendinous arch appeared to be replaced by the venous plexus (Fig. 4D-F).
Thicker nerves tended to be located in the lateral part of the paracolpium, whereas thinner nerves were located in the medial area (Figs. 3, 5). Abundant nerves in the posterosuperior area of the sections (i.e., the space between the distal ureter and the uterine cervix) (Figs. 2A, 4A) appeared to correspond to the so-called detrusor nerves extending toward a site in the bladder at and around the opening of the ureter [27]. In the paracolpium, most of the nerves were cut transversely in the present sections, and thus appeared to run mostly along the postero-anterior axis. Because 2-5 ganglion cell clusters were seen in the paracopium in each of the sections (Figs. 3, 5), the paracolpium appeared to contain the distal or peripheral part of the pelvic autonomic nerve plexus and its branches such as the cavernous nerve, the nerves to the urethra, and the nerves to the internal anal sphincter (NIAS). The plexus and its distal branches were sandwiched by the pubocervical fascia and the rectovaginal septum. The paravaginal ganglion cells and nerves were composed of a mixture of sympathetic and parasympathetic nerve elements (Fig. 5D-F), but the ratio varied significantly between sites: TH-positive sympathetic nerve ganglion cells tended to be dominant in the superolateral part, while nNOS- and/or VIP-positive parasympathetic cells were distributed widely. Most of the nNOS-positive ganglion cells were also positive for VIP (i.e., double positive) (Fig. 3D, E). The NIAS ran postero-inferiorly along the superior fascia of the levator to reach the longitudinal muscle layer of the rectum (Figs. 3A, 5C). The longitudinal course of the cavernous and urethral nerves was seen to originate from the paracolpium and run alongside the urethra in 2 specimens (both, multiparous) (Fig. 6). In the 5 specimens with a thick venous plexus, nerves in the paracolpium were divided into the superior and inferior clusters (Fig. 5C); the superior group appeared to contain candidates for the cavernous and urethral nerves, while the inferior group issued the NIAS. Conversely, few thin nerves were evident between veins. Finally, in the relatively inferior or distal planes we demonstrated, branches (tributaries) of the superior rectal artery (vein) were accompanied by a few thin nerves in the mesorectum behind the rectovaginal septum.
Discussion
The rectovaginal septum (Denonvilliers' fascia) as well as the pubocervical fascia is a well-known gynecologic landmark. The rectovaginal septum has been a target of histological studies [28, 29]. However, demonstration of both fasciae in a single histological section has been rare: Tamakawa et al. [21] did so, but they did not use the common terminology. Hirata et al. [11, 30] noted both of the fasciae, but these were beyond the scope of their study. Our present observations of nulliparous women showed that each of the fasciae was independently connected with the superior fascia of the levator ani. A number of researchers have considered the superior fascia of the levator ani to be the so-called "endopelvic fascia" (reviewed by Hirata et al. [30]), but in the present study we did not use this term because there are many different opinions regarding terminology. Although the small number of specimens we examined was a major limitation of the present study, nulliparous women are likely to exhibit the ideal fascial connection at the tendineous arch of the pelvic fasciae, as proposed by DeLancey [1] (Fig. 1). Destruction of the fascial connection is not always evident even in multiparous women, as Hirata et al. [11] have demonstrated. Thus, the tendinous arch seemed to be more resistant to vaginal delivery than the uterosacral ligament, which is histologically fragmented in multiparous women [11]. Moreover, in multiparous women, the fascial connection seemed to be replaced by the venous plexus, and the veins were embedded in elastic fiber-rich, loose paravaginal tissues. There fore, we hypothesized that, due to vaginal delivery and aging, the pelvic plexus is likely to change from a sheet-like configuration to several bundles in combination with destruction of the basic fascial configuration.
Ball et al. [9] stated that the paravaginal nerves are not localized, but "unfortunately" distributed evenly, unlike the male neurovascular bundles at the 5 and 7 o'clock positions in the prostate. This description is consistent with our observations in nulliparous women. Fig. 7A displays a sheet-like configuration comprising almost evenly distributed paravaginal nerves between the fasciae. In multiparous women with a well developed venous plexus in the paracolpium, veins separated the nerves into superior and inferior groups, and thus the paravaginal nerves tended to be bundled into several fasciculi, although Fig. 7B does not demonstrate this feature well because of technical imperfection. Irrespective of whether they were evenly distributed or bundled, the paravaginal nerves contained both sympathetic and parasympathetic elements. Using fetuses, Yucel et al. [12] demonstrated that the proximal course the cavernous course with nNOS reactivity is close to the mid-urethra at the 5 and 7 o'clock positions, and that at the level of the proximal urethra, it joins the paravaginal nerves at the 2 and 10 o'clock positions of the vagina. In the evenly distributed paravaginal nerves, the female cavernous nerve is likely to localize at a specific site. Nevertheless, demonstration of the longitudinal course was difficult, and we were able to do it in only 2 specimens.
The NIAS (nerves supplying the internal anal sphincter) is another important distal branch of the pelvic plexus although it is not widely known than the cavernous nerves. Because the nerves ran along the posterior or inferior side of the paravaginal venous plexus in multiparous women, tension-free vaginal mesh techniques for uterine prolapse are likely to injure it; this status is similar to that of the levator ani nerve [31]. In fact, the levator ani nerve lies very close the NIAS, but the former (or latter) is located laterally (or medially) to the superior fascia of the levator ani muscle (unpublished data). In the relatively inferior or distal planes we demonstrated, branches (tributaries) of the superior rectal artery (vein) did not accompany abundant nerves in the anterior part of the mesorectum. Conversely, in the same planes or those nearby, the paracolpium was located close to the rectal wall, and the superior fascia of the levator ani was rich in smooth muscles. The latter muscles were continuous with the longitudinal muscle coat of the anal canal, as demonstrated by Arakawa et al. [32]. Thus, the so-called lateral ligament of the rectum seemed to connect with the paracolpium or correspond to part of the latter, although it has been suggested that the ligament may be a surgical artifact [33]. The middle rectal vein, i.e., the major and consistent component of the lateral ligament [34], is likely to drain into the paravaginal venous plexus. Recent macroscopic studies of the lateral ligament of the rectum [7, 35, 36] have indicated that it contains nerve bundles. We consider the NIAS to be a "core" nerve of the lateral ligament.


 XML Download
XML Download