

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
It is known that breast cancer is the most common cancer for women worldwide, and accounts for approximately 25% of all female malignancies with a higher prevalence in developed countries. Breast cancer is the second leading cause of cancer-related death among females in the world [1]. Curcumin (1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) (Figure 1), which is extracted from the plant Curcuma longa, is an antioxidant that exerts antiproliferative and apoptotic effects. The expressions and activities of various proteins, such as inflammatory cytokines and enzymes, transcription factors, and gene-products linked with cell survivals and proliferation, can be modified by curcumin [2]. Since curcumin possesses anti-inflammatory, antioxidant, and antitumoral effects, it has been studied vigorously as a chemopreventative agent in some cancer models and used in the therapeutic arsenal in clinical oncology. However, curcumin is insoluble and instable in water. The solubility of curcumin could be enhanced by utilizing the solubilizing properties of rubusoside. In addition, the selective delivery of synthetic analogs or nanotechnology-based formulations of curcumin to tumors may improve the chemopreventive and chemotherapeutic effects. The focus of this short review is to describe how curcumin participates in the regulation of oncogene protein expression in breast cancer cells.
THE EFFECT OF CURCUMIN ON ONCOGENE PROTEIN EXPRESSION
The effect of curcumin on the oncogene proteins of proliferation and apoptosis
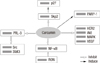
Curcumin inhibits the proliferation of various tumor cells in culture and prevents carcinogen-induced cancers in rodents. In xenotransplant or orthotransplant animal models, the growth of human tumors was inhibited by curcumin alone or in combination with chemotherapeutic agents or radiation [2]. Banerjee et al. [3] reported that curcumin induced G2/M arrest and apoptosis, inhibited cell proliferation by inhibiting the assembly dynamics of microtubules, and further activated the mitotic checkpoint in MCF-7 cells. Furthermore, cells were accumulated in the G(1) phase of the cell cycle, and curcumin suppressed the expression of zeste homolog 2 (EZH2) gene via the stimulation of three major members of the mitogen-activated protein kinase (MAPK) pathway: c-Jun NH2-terminal kinase (JNK), extracellular signal-regulated kinase (ERK), and p38 kinase (Figure 2) [4].
The apoptosis is involved in the curcumin-induced inhibition of tumor cell growth. It was found that curcumin inhibited the expression of Ki-67, proiferating cell nuclear antigen (PCNA), and p53 mRNAs in breast cancer cells, and induced Bax mRNA expression with the down-regulation of p21 mRNA in the human mammary epithelial cell line (Figure 2) [5]. The development of breast cancer is associated with the abnormal activation of the Wnt/β-catenin signaling pathway, and curcumin inhibits the expression of β-catenin, cyclin D1, and slug in both MCF-7 and MDA-MB-231 cells (Figure 2) [6].
Maspin is a serine protease inhibitor, which suppresses tumor growth and metastasis in vivo and tumor cell motility and invasion in vitro. The results of Prasad et al. [7] indicated that the maspin gene expression was increased by curcumin in MCF-7 cells with the up-regulation of p53 protein and down-regulation of Bcl-2 (Figure 2). Another report indicated that curcumin reduced the expression of Bcl-2 by up-regulating the expression of miR-15a and miR-16 in MCF-7 cells (Figure 2) [8]. However, the data of Chiu and Su [9] showed that curcumin inhibited the proliferation of MDA-MB-231 cells either through up-regulating p21 expression or up-regulating the Bax to Bcl-2 ratio. Moreover, the AIP-1/Alix protein, which is an inhibitor protein of paraptosis, was progressively down-regulated in curcumin-treated malignant breast cancer cells (Figure 2) [10]. In curcumin-induced paraptosis, mitochondrial superoxide acted as a critical early signal and proteasomal dysfunction was mainly responsible for the paraptotic changes associated with estrogen receptor (ER) dilation [10]. It was also found that curcumin inhibited breast cancer cell motility and invasion by directly inhibiting the function of α6β4 integrin [11].
The effect of curcumin on nuclear factor-κB
Most anticancer agents activate nuclear factor-κB (NF-κB), which mediates cell survival, proliferation, and metastasis. It has been found that curcumin inhibits the migratory activity of breast cancer cells, proliferative rate, adhesion, and invasion through down-regulating the expression of NF-κBp65 [9,12-14]. Human epidermal growth factor 2 (HER2) is an important oncoprotein, which is overexpressed in about 15% to 25% of breast cancers. Curcumin decreased HER2 oncoprotein, phosphorylation of Akt, MAPK, and the expression of NF-κB in both BT-474 and SK-BR-3-hr cells (Figure 3) [15]. Furthermore, curcumin enhanced the efficacy of chemotherapy by tailoring p65NF-κB-p300 cross-talk in favor of p53-p300 for breast cancer [16]. Curcumin also shows a potential therapeutic role in blocking recepteur d'origine nantais (RON) tyrosine kinase-mediated invasion of carcinoma cells by affecting p65 protein expression and transcriptional activity via NF-κB (Figure 3) [17].
The effect of curcumin on vascular endothelial growth factor
Curcumin may provide a clinically useful tool for the suppression of the elaboration of vascular endothelial growth factor (VEGF) in tumor cells (Figure 3). It has been found that curcumin suppressed breast tumor angiogenesis by abrogating osteopontin or medroxyprogesterone acetate induced VEGF expression [18,19]. Curcumin inhibited α6β4 signaling and functions by altering intracellular localization of α6β4, and prevented its association with signaling receptors such as the epidermal growth factor receptor (EGFR) and Akt [20,21]. In addition, the combination of epigallocatechin gallate (EGCG) and curcumin is efficacious in both in vitro and in vivo models of ERα-breast cancer. In this processes, the regulation of VEGFR-1 may play a key role in the antitumor activities [21].
The effect of curcumin on other oncogenes
Curcumin has been studied vigorously as a chemopreventative in several cancer models. Since cleaved poly (ADP-ribose) polymerase 1 (PARP-1) plays a significant role on cellular protection against radiation and estrogen-induced DNA damage mediated by curcumin, targeting PARP-1 may provide an effective way of maximizing the therapeutic value of antioxidants for cancer prevention (Figure 3) [22]. On the other hand, the F-box protein S-phase kinase-associated protein 2 (Skp2), which acts as an oncogene through targeting p27 for degradation, is overexpressed in many different human cancers. Because curcumin induces p27 expression and growth arrest through the inhibition of Skp2 in MDA-MB-231 cells, the therapeutic strategies designed to reduce Skp2 may therefore play an important clinical role in treatment of ER/HER2 negative breast cancers (Figure 3) [23]. In addition, phosphatase of regenerating liver-3 (PRL-3) was also treated as a potential target for anticancer drugs based on its involvement in tumor metastasis. It was found that curcumin inhibited the phosphorylation of Src and stat3 partly through PRL-3 down-regulation, raising its possibilities in therapeutic regimen against malignant tumor (Figure 3) [24].
THE EFFECT OF CURCUMIN AND MITOMYCIN C COMBINATION TREATMENT ON BREAST CANCER CELLS
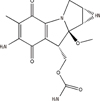
Mitomycin C (MMC) (Figure 4), a potent DNA cross-linker and antineoplastic agent, is usually used to fight various cancers. However, the use of MMC is limited because the prolonged use of MMC will result in permanent kidney or bone marrow damage and secondary tumors in normal cells. It has been found that curcumin improves MMC-based chemotherapy by simultaneously sensitizing cancer cells to MMC and reducing MMC-associated side-effects, increasing cell viability, and further decreasing lipid peroxidation and DNA damage [25,26]. The combination treatment of MMC and curcumin reduces the toxic effect of MMC by inhibiting glucose regulatory protein (GRP58)-mediated DNA cross-linking through the ERK/p38 MAPK pathway (Figure 5) [27]. Another report indicated that curcumin enhanced antiproliferative effect of MMC in human breast cancer MCF-7 cells via the p38 MAPK pathway [25]. The cell cycle arrest was associated with the inhibition of cyclin D1, cyclin E, cyclin A, cyclin-dependent kinase 2 (CDK2), and CDK4. But the cell cycle inhibitors p21 and p27 were induced in MCF-7 cells and MCF-7 xenografts (Figure 5) [25].
ENHANCEMENT THE SOLUBILITY AND STABILITY OF CURCUMIN
Though curcumin has been indicated as highly cytotoxic towards various cancer cell lines, its insolubility and instability in water contributes to low bioavailability. On the other hand, photodegradation and low bioavailability are major hurdles for the therapeutic use of curcumin. However, the solubility of curcumin could be enhanced by utilizing the solubilizing properties of rubusoside, and the rubusoside-solubilized curcumin successfully inhibited cell viability in human colon, breast, and pancreatic cancer cell lines [28]. In order to increase curcumin photostability and enhance its anticancer activity against MCF-7 breast cancer cells, Mulik et al. [29] formulated the transferrin-mediated solid lipid nanoparticles (Tf-C-SLN), which enhances the anticancer effect of curcumin in breast cancer cells in vitro. Moreover, it was found that curcumin conjugated with phosphatidylcholine increased curcumin bioavailability five-fold compared to original curcumin [30]. The prodrugs, which are produced by mono-PEGylation of curcumin, are stable in buffer at physiological pH and released curcumin readily in human plasma [31].
The polycurcumins have high drug loading efficiency and can be used as backbone-type conjugates. The polycurcumins could fix drug loading contents, stabilize curcumin in their backbones, and tailor water-solubility. Tang et al. [32] made the high molecular weight curcumin polymers (polycurcumins) through condensation polymerization of curcumin. It was found that the polyacetal-based polycurcumin was not only highly cytotoxic to MCF-7 breast cancer cell lines but also showed significant antitumor activities in SKOV-3 intraperitoneal xenograft tumor models [32].
CONCLUSION
Recently, the potential effect of curcumin on cancer cells has been recognized by the scientific community in the world, and the molecular biological approaches help to elucidate the underlying mechanisms of actions on curcumin in tumor cells. However, the molecular mechanisms underlying the antitumor activity of curcumin have not been very clear until now. Another thing is that the molecular mechanism of curcumin on tumor cells was usually studied with tumor cell lines in vitro, and the molecular mechanisms in vivo need to be further investigated. More sophisticated technologies will have to be applied in conjunction so that curcumin derivatives could be used for rational cancer therapy.

 XML Download
XML Download