

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Bartter syndrome (BS) is an autosomal recessive inherited disorder characterized by hypokalemic metabolic alkalosis with normal or low blood pressure despite hyperreninemic hyperaldosteronism (1, 2). BS is clinically classified into antenatal or neonatal BS (aBS) and classic BS (cBS) as well as five subtypes based on the underlying mutant gene, all of which are expressed in the tubular epithelial cells of the thick ascending limb of the loop of Henle (3-8). Specifically, BS type I (BS I) is caused by mutations in the SLC12A1 gene encoding the apical sodium-potassium-chloride co-transporter, NKCC2 (3). BS II is caused by mutations in the KCNJ1 gene encoding the apical inwardly rectifying potassium channel, ROMK (4). BS III is caused by mutations in the CLCNKB gene encoding the basolateral chloride channel, ClC-Kb (5, 6). BS IV is characterized by mutations in the BSND gene encoding barttin, the β-subunit for ClC-Ka and ClC-Kb (7). Finally, BS V is characterized by gain-of-function mutations in the CASR gene encoding the basolateral calcium-sensing receptor, CaSR (8). Unfortunately, for BS there is presently no curative treatment (1, 2). Therapy of BS is today accomplished by the correction of hypokalemia and the use of prostaglandin synthetase inhibitors. However, lack of rigorous therapeutic control may lead to progression to chronic renal failure. In a previous study, we found that BS III was the most common genotype (23 of 26 patients) in Korean children with BS and a nonsense mutation of p.W610X was the most common mutation in CLCNKB (9). This nonsense mutation was detected in 25 of 46 (54.3%) alleles of the patients with BS III, and 18 of 23 (78.3%) patients with BS III harbor p.W610X in one or both alleles (9).
Translational read-through induction is an approach to rescue a full-length protein from a gene with a premature stop codon by modifying gene expression, i.e., reducing the accuracy of translation elongation and the efficacy of the translation termination machinery (10). Several studies have been tried translational read-through induction as a novel therapeutic approach for various kinds of diseases caused by nonsense mutations using aminoglycosides or their derivatives, well-known pharmacological agents that can induce ribosomal read-through (11-16). While mis-incorporation of an amino acid at a stop codon generally occurs with a frequency of 10-4 in intact cells under normal conditions (11), the frequency increases with a readthrough efficiency of up to 1% to 25% in the presence of aminoglycosides (10). Aminoglycosides bind to the 18S ribosomal RNA and induce a conformational change in its decoding site, thereby inducing a reduction in proofreading, an induction of near-cognate aminoacyl-tRNA mis-incorporation, and translation of full-length, functional proteins despite the presence of nonsense mutations (10, 17). Additionally, once read-through efficiency exceeds 0.5%, nonsense-mediated mRNA decay is significantly reduced with further promotion of read-through (10). This effect may prove significant in recessive disorders resulting from nonsense mutations, in which proteins are rarely expressed (10). In such cases, even 1% of normal protein function may restore a clinically less severe or close to normal phenotype (10). It has been demonstrated that aminoglycosides can induce a read-through of nonsense mutations with an efficiency of up to 20% in various genetic disorders (18-20).
In this study, we tried in vitro translational read-through induction of the p.W610X mutant CLCNKB, the most common CLCNKB mutation in Korean patients with BS, using G418 (geneticin), a neomycin derivative aminoglycoside.
MATERIALS AND METHODS
CLCNKB cDNA cloning and site-directed mutagenesis
We purchased a wild-type full-length human CLCNKB cDNA (GenBank NM_000085), which was cloned into pcDNA3.1 (+) (Invitrogen, Carlsbad, CA, USA) using HindIII and XbaI cloning sites, from Source BioScience (Nottingham, UK). The entire length of inserted cDNA except for the 3' stop codon were amplified by polymerase chain reaction (PCR) and re-cloned into pcDNA3.1D/V5-His TOPO (Invitrogen). This recombinant vectors can express C-terminal V5-His tagged recombinant ClCKb protein (ClC-Kb-V5-His). V5 and His were tagged for the convenience of the immunochemical detection of the protein in the later steps. Site-directed mutagenesis (c.1830G > A in CLCNKB) was generated using the PCR-based DpnI-treatment method. This mutagenesis was confirmed by bidirectional sequencing.
Cell culture and transfection
Twenty-four hours prior to transfection, MDCK cells (canine kidney epithelial cells) were seed in 12-well plates or 2-well Lab-Tek chamber slides (80,000 cells per well or chamber with 2 mL of Dulbecco's Modified Eagle's Medium (DMEM). The cells were incubated at 37℃ in an atmosphere of 5% CO2. After 24 hr of incubation, when the cells were 50%-70% confluent, the polarized MDCK cells were transfected with purified plasmids (containing either wild-type or p.W610X mutant CLCNKB) using jet-PEI™ (Polyplus transfection, Illkirch, France) in accordance with the manufacturer's protocols. Transfection was conducted for 8 hr with 2 µg of DNA and 4 µL of jetPEI™ reagent in 150 mM NaCl in fresh media. Co-transfection with pcDNA3.1/NT-GFP-TOPO vector (Invitrogen) was carried out in order to assess transfection efficacy.
G418 treatment for read-through induction
After 8 hr of transfection of polarized MDCK cells, the culture media were changed into fresh DMEM containing 0, 75 or 150 µg/mL of G418 (geneticin, Roche Applied Science, Indianapolis, IN, USA). Media with/without G418 were changed every 24 hr and after 60 hr, and the transfected MDCK cells were harvested and analyzed. We divided the samples into 4 groups; Group 1, cells transfected with wild-type CLCNKB without G418 treatment; Groups 2, cells transfected with p.W610X mutant CLCNKB without G418 treatment; Group 3, cells transfected with mutant plasmid with 75 µg/mL of G418; Group 4, cells transfected with mutant plasmid with 150 µg/mL of G418. Non-transfected MDCK cells were employed as a control.
Immunohistochemistry
Each group of cells in 2-well Lab-Tek chamber slides was fixed for 20 min in PBS containing 4% paraformaldehyde. After washing three additional times with PBS, the cells were permeabilized with 0.1% Triton X-100 for 1 hr and blocked in PBS containing 1% BSA and 0.1% Triton X-100 for 1 hr. After 60 min of incubation with 1:200 diluted His-probe (H-3) (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at room temperature, and the cells were washed three times in PBS and incubated for 30 min with 1:1,000 diluted Alexa Fluor 594-labeled goat antimouse IgG (Invitrogen) at room temperature.
The prepared cover slips were mounted onto slides with DAPI (Invitrogen) for nuclear staining. The stained cells were subsequently analyzed via a confocal laser microscopy (LSM 510 META; Carl Zeiss, Jena, Germany). To evaluate protein expression, 5 stained fields were examined, and the percentage of cells stained along plasma membrane was assessed. Total cell count examined was from 150 to 200.
Western blot
Each group of cells in 12-well culture plates were rinsed twice with PBS and lysed via solubilization in lysis buffer (150 mM/L NaCl, 50 mM/L Tris, pH 7.5, and 0.2% NP40). Lysates were centrifuged at 4℃ to remove insoluble matter. The whole cell lysates were prepared for electrophoresis by addition of sample buffer. Additionally, for fraction of plasma membrane proteins from cell culture samples, Qproteome plamsa membrane protein kit (Qiagen, Hilden, Germany) was used. A total protein sample were separated on 8% SDS-polyacrylamide gel and transferred onto nitrocellulose membrane in transfer buffer. Nonspecific binding sites were blocked by incubation with blocking solution for 1 hr at room temperature, after 1 hr incubation with anti-V5-HRP antibody (Invitrogen). All antibodies were used at concentrations recommended by the manufacturer. Immunoreactive bands were visualized using enhanced chemiluminescence.
RESULTS
Immunohistochemistry
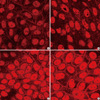
While MDCK cells transfected with wild-type CLCNKB plasmid (Group 1) expressed ClC-Kb-V5-His protein along the plasma membranes (Fig. 1A), cells transfected with mutant plasmid (Group 2) did not express the protein (Fig. 1B). However, after G418 treatment, mutant plasmid transfected cells expressed the fusion protein along the plasma membranes, and the degree of protein expression revealed no significant difference between 75 µg/mL (Group 3, Fig. 1C) and 150 µg/mL (Group 4, Fig. 1D) of G418 treatment. The percentage of cells stained along plasma membrane was 63% and 2% in MDCK cells transfected with wild-type CLCNKB plasmid and in cells transfected with mutant plasmid, respectively (Fig. 1A, B). The mutant plasmid transfected cells with low dose G418 and the cells with high dose G418 expressed the protein in 27% and 33% cells, respectively (Fig. 1C, D). The confocal laser microscopic examination showed that the membrane expression of the protein was localized to the basolateral membranes (Fig. 2B). Arrows indicate the protein that was basolaterally stained (Fig. 2B).
DISCUSSION
In the present study, we confirmed immunohistochemically that G418 treatment induced translational read-through induction of the p.W610X mutant cDNA in polarized MDCK cells. In addition, the translated proteins were transported properly to the basolateral plasma membranes. Although the residue that was substituted for the UGA stop codon remained to be elucidated in this study, the most frequently reported substitution for the UGA stop codon was UGG, which encoded for tryptophan (14, 21). As a general rule, glutamine is inserted at the nonsense UAG or UAA read-through sites, whereas UGA sites miscode to tryptophan (14, 21). This result is encouraging in terms of the possible treatment of aminoglycosides in BS patients harboring the p.W610X mutation in the aspect of gene specificity during the induction of read-through.
In this study, western blot analysis was performed to determine the effect of G418 on the levels of protein expression for wild type, mutant without G418, and mutant with G418. However, western blot analysis of any groups of cells showed no clearly recognized bands. It is possible that the transfection efficacy of the cells was not high enough to get a target band by western blot analysis. Co-transfection with pcDNA3.1/NT-GFP-TOPO vector in this study revealed that the transfection efficacy was less than 20% with GFP and DAPI staining (data not shown). Further studies to determine the read-through effect on the levels of mRNA or protein are needed.
In this study, it is difficult to measure the efficacy of the readthrough. Because several factors including the kind of stop codon, and the identity of the fourth nucleotide have been recognized with regard to the efficiency of read-through induction, the efficacy of this study may be estimated according to these rules (22). For example, the efficacy of read-through in UGA stop codon is superior to that in UAG or UAA. Another study reported that G418 suppressed a UGA stop codon with different efficiencies dependent on the fourth nucleotide in order of C > A, G > U (23). In case of CLCNKB p.W610X mutation, the stop codon is UGA and the fourth nucleotide is G, which suggested intermediate or higher efficiency. The reason for this preference remains unclear, but may relate to the ability of the aminoglycoside to allow some mismatch between certain codon and anticodon pairings, with the UGA mismatch being more permissive than either UAG or UAA (16, 23).
Although we have shown for the first time that aminoglycoside induced-ribosomal read-through induces the production of protein in cells expressing mutant CLCNKB p.W610X by immunohistochemistry, this study has some limitations and additional experiments are needed. For example, if the protein was detectable by immunoblot analysis, the rate of induction of protein expression following G418 treatment of MDCK cells can be determined. Additionally, since the effect of G418 is medicated intracellularly, assay to measure intracellular G418 content in MCDK cells may be helpful. Finally, functional studies to determine whether translated protein function as chloride channel are necessary to explore the usefulness of aminoglycoside for the treatment of BS.
We use G418 because of the best termination suppression activity in vitro, however, these is a risk of side effects to use aminoglycosides clinically (11, 23-25). The chemical composition of aminoglycoside may influence on the efficiency of read-through and specific side effects (16, 23). For example, it is known that G418 may elicit specific side effects, such as vagina and oral mucosa ulceration in dogs (26). To improve termination-suppression activity and to lower the toxicity, several newer drugs have recently been identified (16, 27). NB 54, for example, is a newly derived aminoglycoside tailored specifically for the suppression of nonsense mutations and evidences a suppression effect several-fold greater than that of gentamicin (16, 27). This drug also evidences far lower toxicity than gentamicin or amikacin, thereby increasing its potential for chronic therapeutic use (27). Another excellent example is PTC124, which is an orally bioavailable small organic molecule with no antibiotic properties (28). This drug can specifically promote the read-through of disease-causing premature termination codons but not the termination of normal stop codons (28). Additionally, PTC124 has been associated with no serious adverse side effects (28). It is necessary to determine whether either of these new drugs will facilitate the effective read-through of nonsense mutations of CLCNKB.
In conclusion, our study shows that the p.W610X mutation in CLCNKB can be another candidate for trial of translational read-through induction as a therapeutic modality. Further studies to determine rate of induction of protein expression induced by G418, effect of G418 on CLCNKB p.W610X message level, analysis of intracellular G418 content and functional studies are required prior to the final clinical trial.

 XML Download
XML Download