

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Inflammatory bowel disease (IBD) is defined as chronic inflammation of the bowel of unknown etiology and normally refers to Crohn's disease (CD) and ulcerative colitis. While specific causes are unknown, it is thought to be associated with the bowel's immunological function, especially activated T-lymphocytes.(1,2) T-lymphocytes secrete cytokines which regulate immune response and according to their function, are divided into T-helper 1 (Th1) (IL-2, interferon-γ; regulation of cell-mediated immunity) and T-helper 2 (Th2) (IL-4, IL-5, IL-10, IL-13; regulation of humoral immunity) cells.(3,4) Dysregulation of T-lymphocytes in IBD leads to imbalance of these cytokines, causing an influx of inflammatory cells such as T-lymphocytes and macrophages and subsequent tissue damage.(5) Also, genetic and psychological susceptibility, altered bowel microflora and various other factors are thought to be involved in the inflammatory process.(6,7) IBD progresses either gradually over a long period or acutely over several days.(8) With lack of a definitive treatment option due to an ill-defined etiology, most patients enter a chronic disease state.(2)
Current medical treatment for IBD mainly consists of pharmacological intervention. Rather than achieving a cure, the current aim of treatment is to improve quality of life by inducing remission, preventing recurrence and controlling the symptoms.(9) Drugs used include aminosalicylates (sulfasalazine, mesalamine), corticosteroid-based anti-inflammatories, antibiotics (metronidazole), immune boosters (levamisole, Bacillus Calmette Guerin (BCG)), a mast cell stabilizer (cromolyn sodium), and allopathic medications including antidiarrheals, antispasmodics and cholestyramine.(9,10) An immunosuppressant (azathioprine, mercaptopurine, methotrexate, cyclosporine) can be used when other drug treatments are ineffective.(10) However, many of these drugs face limited use because of ineffectiveness and adverse side effects.
Due to issues facing drug treatments, treatment using mesenchymal stem cells (MSC) is surfacing as a new method of treatment. MSCs are multipotent non-hematopoietic progenitor cells from adult bone marrow which can differentiate into various cells that make up fat, cartilage, bone, muscle, stroma, nerves, etc.(11) Currently, clinical research is aimed towards achieving tissue regeneration with MSCs.
MSCs have a low expression of antigens such as HLA-DR, and therefore produce less adverse immune responses when injected in vivo.(12,13) MSCs can be genetically modified, and they can produce various growth factors or cytokines continuously or for certain periods of time when injected.(14) They cannot present antigen due to absent MHC-II and co-stimulatory molecules, but they play an important role in proliferation and growth of hematopoietic stem cells and produce various cytokines and regulatory molecules that regulate granulocyte, macrophage and dendritic cell production, hemopoiesis and immune response.(11,12)
MSCs with low immunogenicity, immunomodulatory function, and ability to differentiate into various cell lineages have not shown graft versus host disease or caused damage to the recipient's immune system.(15,16) Also, by forming chimeras, they have led to donor-specific immune tolerance.(17)
Cytotherapy using stem cells is receiving attention as the next generation treatment method as it can be used in rejection or incurable illnesses that current medicines cannot solve.(12) Cytotherapy also can be applied to various fields of medicine including cardiovascular, neurological, hematological, and immunological systems, and also hereditary, hepatic and endocrine diseases. Utilization of MSCs in the treatment of IBD requires more in-depth research to clarify its mechanism of action. Therefore, in the present study, we examined the effect of MSCs in the murine model of TNBS-induced colitis.
METHODS
1) Induction of colitis and treatments
Female 8-week-old BALB/c mice (weight range 20~25 g) were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). All mice were housed in specific pathogen-free conditions and in accordance with institutional guidelines. Colitis was induced by rectal administration of 2 mg of 2,4,6-trinitrobenzene sulfonic acid (TNBS, Sigma Chemical Co., St. Louis, MO, USA) in 35% ethanol (Merck, Germany) using a vinyl catheter positioned 4 cm from the anus (20 mice per group). After instillation they were kept vertically for 60 seconds with mice faced down for effective absorption and prevention of regurgitation of administered TNBS or ethanol. The MSC treatment group was treated with 5×105 cells/150 µl in Dulbecco's Phosphate Buffered Saline (DPBS, GIBCO, NY, USA) isolated from the mice inguinal fat pad, via tail vein 24 hrs after colitis induction. The control group was treated with 35% ethanol (Et-OH).
2) Assessment of inflammation and histology
Mice were weighed daily. Body weight loss was determined by percentage of weight loss from baseline body weight. Mice were sacrificed on each 3 (n=10) and 10 days (n=10) after the TNBS administration. The longitudinally divided colons were rolled up and fixed in 4% buffered formalin. Fixed tissues were embedded in paraffin, and 4~6 µm sections were stained with H&E for histologic grading. An experienced pathologist blinded to treatment allocation scored all sections: (1) percentage of colon involved, (2) infiltration of mononuclear cells, and (3) alteration & erosion. The severity was scored on a scale of 0~4 as follows: 0, absent; 1, weak; 2, moderate; 3, severe; 4, most severe. Grading was done in a blinded fashion by one of the authors.
3) Flow cytometric analysis and cytokine assay
The surface molecules of T cells were observed in the peripheral blood and spleen by the use of mAb directed to CD4, CD8a, CD19, CD25 and a control isotype conjugated with fluorochromes (FITC, PE). All Abs were purchased from BD PharMingen (San Diego, CA, USA) and analysis of the samples was carried out on a FACS Vantage flow cytometer (BD Biosciences, USA). Splenocytes were cultured in 200 µl of RPMI 1640 medium supplemented with L-glutamine, antibiotics, 10% FBS and 2 µg/ml anti-CD28 mAb in 96-well plates (Costar, USA) precoated with 5 µg/ml anti-CD3 mAb for 72 hrs. Cytokine concentrations in the culture supernatants were determined by specific ELISA according to the manufacturer's instructions (R&D system, USA).
RESULTS
1) Measurement of survival rate and weight loss
To investigate the role of MSC in experimental colitis and the therapeutic effect of MSC, colitis was induced in BALB/c mice. Colitis was induced by intrarectal administration of TNBS, which haptenates autologous colonic proteins with trinitrophenol. Administration of TNBS resulted in diarrhea and wasting disease with an increased mortality rate of approximately 60% after 10 days (Fig. 1). TNBS colitic mice treated with MSCs showed a markedly increased survival rate of 90% compared to TNBS colitic mice of 40% (Fig. 1).
Similar decrease in body weight was observed in both TNBS colitic mice and MSC treated colitic mice at day 1 (Fig. 2). Decrease in body weight (maximum of more than 10%) was observed until day 3 followed by gradual recovery in TNBS colitic mice (Fig. 2). However, MSC treated colitic mice showed no further weight loss after day 1 and showed rapid weight recovery (Fig. 2).
Thus, MSC treatment in colitis-induced mice resulted in increased survival rate and decreased weight loss.
3) Flow cytometric analysis
Induction of TNBS colitis resulted in a slight decrease in CD4+ T cells in the blood and spleen (except day 10 in spleen), whereas MSC treatment showed increase in it. CD8a+ T cells showed a slight decrease after TNBS treatment in both blood and spleen samples, but increased after MSC treatment. CD19+ T cells in the spleen showed a 30~40% decrease in the TNBS group compared to control/Et-OH group, but showed a dramatic decrease after MSC treatment, and CD4+CD25+ T cells showed a marked increase in TNBS/MSC group compared to TNBS group in the blood on day 3 (Table 2).
4) Measurement of cytokine production in culture medium
In order to determine the treatment effect of MSC in TNBS colitis, splenocytes producing various cytokines were cultured and ELISA was performed.
Concentrations of IFN-γ and IL-12 from the Th1 group (IFN-γ, TNF-α, IL-12) were significantly lower in the MSC treated group compared to the control (Et-OH) and TNBS groups, and concentrations of TNF-α were significantly higher in the MSC treated group compared to the TNBS group on day 3 but markedly decreased by day 10 which showed that there is a decreasing effect on pro-inflammatory markers (Fig. 4).
Concentration of IL-4 in the Th2 group (IL-4, IL-6, IL-10) was significantly lower in the MSC treated group compared to the TNBS group on day 10. In contrast, the concentration of IL-6 was significantly higher in the MSC treated group on day 3. The concentration of IL-10 on day 3 showed no noticeable differences in all 3 groups, but the MSC treated group showed a significant increase on day 10 compared to the TNBS group (Fig. 4).
DISCUSSION
In this study, we examined the effects of MSC on TNBS-induced colitis, which is a CD4+ T-cell mediated autoimmune disease.
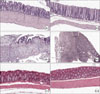
Clinical results from H&E staining showed a significant decrease in both the microscopic inflammatory process and in damage in the TNBS/MSC group compared to the TNBS group. On day 10, all of the involved regions showed a significant decrease in alteration and erosion. Mononuclear cell infiltration was also decreased, but was not statistically significant. These are all signs of a decrease in the inflammatory process, which proves that MSC has an anti-inflammatory effect on microscopic inflammation.
IFN-γ and IL-12 decreased in the TNBS/MSC group compared to the TNBS group, which showed a decrease in Th1 response. IFN-γ and IL-12 both significantly decreased on both days 3 and 10. This is possibly due to termination of the IFN-γ - IL-12 - IFN-γ - IL-12 loop by MSC. In this loop, increase in IFN-γ leads to an increase in IL-12, which itself causes the IFN-γ to increase again in a loop.(18) This illustrates the Th1 polarization (decrease in Th1 response that is responsible for cellular immunity) and inhibition of inflammation that is triggered by the inflammatory IFN-γ cytokine.
IL-4 decreased and IL-10 increased in the TNBS/MSC group compared to the TNBS group, which showed an anti-inflammatory response in the Th2-mediated immune response. IL-4 acts on the T-helper 0 (Th0) cells to differentiate into Th2 cells.(19) Subsequently, IL-4 acts on Th2 cells to produce Th2 cytokines such as IL-5, IL-6, IL-13 and even IL-4 (paracrine effect).(20,21). Therefore, a significant decrease in IL-4 cells attenuates the Th2 immune response, leading to decreased humoral immunity. Also, IL-4 works with IFN-γ, where an increase in IFN-γ also leads to a subsequent increase in IL-4.(19) IL-10 also antagonizes the production of Th1 cytokines IFN-γ and IL-12.(22) Therefore, the significant decrease in IFN-γ correlates with the significant decrease in IL-4 and the significant increase in IL-10, showing that the TNBS/MSC group is effective in attenuating the inflammatory process. Aggarwal and Pittenger(14) also reported a decrease in IFN-γ and an increase in IL-10, but also an increase in IL-4 in their study using human MSC, which showed a Th2 shift. Our current results might suggest a general decrease in both Th1 and 2 inflammatory processes by MSC. In addition, the significant increase in IL-6 level in day 3 TNBS/MSC group is explainable as IL-6 causes increased migration and activation of MSC.(23)
Cell surface markers from the TNBS/MSC group also showed the anti-inflammatory effect of MSC. A significant increase in CD4+CD25+ T cells in the blood sample of the TNBS/MSC group showed that T-regulatory cells that attenuate the inflammatory process increased.(24,25) This is in line with the study by Veltkamp et al.(26), which had shown that CD4+CD25+ T-regulatory cells prevent experimental colitis in the bone marrow transplanted murine model. Also, a significant decrease in CD19 in the spleen sample of the TNBS/MSC group on day 10 showed effective attenuation of the Th2 response as CD19+ B cell subset population blocks T regulatory cells and exacerbates ileitis in murine model of CD.(27,28) However, the general decrease in CD19 level in the TNBS group compared to the control group requires further research.
However, cell surface markers from the TNBS group yielded different results. CD8a (blood, day 4) and CD4 (blood and spleen, day 10) in the TNBS/MSC group all showed a significant increase compared to the TNBS group, which shows that there is still inflammation and a Th1 response. This could have been due to a remaining inflammatory process that was not attenuated by MSCs.
Also, TNF-α levels did not correlate with other cytokine levels. TNF-α levels significantly increased on day 3 in the TNBS/MSC group compared to the TNBS group, which shows that there is a significant inflammatory process present. This result did not correlate with other cytokine levels which all showed a decrease, signifying a decreased inflammatory process. Whereas this study showed no significant decrease in the TNF-α levels by MSCs, Lee et al.(29,30) showed that DSG and anti-4-1 BB mAb leads to a decrease in TNF-α levels. However, TNF-α significantly increased only on day 3 but not on day 10 in this study, which can be interpreted as MSCs working more effectively in the later part of the inflammation process and not in the earlier part. Also, results from H&E staining showed a significant decrease in the microscopic inflammatory process on day 10 which correlates to the lack of a significant rise in these cytokine levels by then.
There is much research being done on the use of MSCs in treating intestinal diseases. It has been shown that adipose-derived MSCs can be used to treat rectovaginal fistulas in perianal CD without any adverse effects.(31) Research has suggested a great potential for the application of MSC. However, the mechanism of action still requires more in depth clarification, which will provide the direction and focus for future study. The current study would have been able to yield more reliable results if the sample sizes were larger. The addition of an apoptosis detection assay to monitor the decrease in specific cell populations would also have given more accurate findings. In addition, monitoring other cytokines such as CD11 (leukocyte adhesion, macrophage levels) would have given more insight into the anti-inflammatory process. Research comparing the effect of using MSC treatment along with conventional treatment versus conventional treatment alone could be meaningful. Because of MSC's immunomodulatory effect, synergy with conventional treatment to increase treatment effectiveness and decrease side effects can be expected.
In conclusion, MSC's immunomodulatory effect in itself was shown to have a possible therapeutic effect in the murine IBD model, especially in CD. With further research, it could be possible to utilize MSC in the clinical treatment of IBD if subsequent experiments can prove the safety of MSC therapy.





 XML Download
XML Download