

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Introduction
Early vaccination strategies used live or attenuated pathogens to induce adaptive immunity [1]. However, the re-activation of attenuated vaccine materials and the possible tumorigenicity of inactivated oncogenic viruses are important safety concerns [2]. Thus, non-pathogenic materials including inactivated toxins, synthetic peptides, and recombinant subunit proteins have been considered as vaccine materials [3]. However, these antigens are poor in immunogenicity and must be given with supplemental materials to potentiate the vaccination capacity [3]. Such supplemental materials are termed adjuvants and can be divided into two classes depending on their mode of action: efficient delivery of vaccine materials and/or stimulation of the immune system (Tables 1, 2) [1]. Adjuvants facilitating vaccine delivery include liposomes, nanogels, oil-in-water emulsions, and virosomes targeting the co-administered antigens to professional antigen-presenting cells (APCs) [4]. Adjuvants that stimulate the immune system include molecules binding to intracellular receptors including Toll-like receptors (TLRs), Nod-like receptors, and RIG-I–like receptors and to cytosolic DNA sensors, all of which modulate the immune response [1]. However, most adjuvants have been evaluated in the context of parenteral immunization; thus, it is not clear how well the adjuvants function in the mucosal immune compartment.
The Mucosal Immune System
Mucosal surfaces cover 400 m2 of the body including the gastrointestinal, urogenital, and respiratory tracts [5]. Mucosae are continuously exposed to microbiota and antigens. The gastrointestinal mucosa is especially prone to the development of tolerogenic microenvironments, where luminal antigens may persist. The mucosal immune system has both inductive and effector sites differing in terms of their anatomical and functional characteristics [6]. The major mucosal immune inductive sites include gut-associated lymphoid tissue (GALT) and the nasopharyngeal-associated lymphoid tissue (NALT). GALT includes Peyer's patches, mesenteric lymph nodes, and isolated lymphoid follicles, while NALT includes tonsils/adenoids, inducible bronchus-associated lymphoid tissue, cervical lymph nodes, and hilar lymph nodes. Mucosal immune inductive sites are covered by follicle-associated epithelium (FAE), which is composed of enterocytes and M cells.
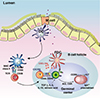
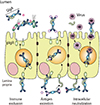
M cells are specialized epithelial cells for antigen uptake [7]. These cells are overlaid by a thin mucus layer and possess short irregular microvilli [8]. M cells can transfer antigens via transcytosis to APCs located in pockets within M cell clusters [9]. Dendritic cells that come in contact with antigens transcytosed through M cells enter the interfollicular T cell zone to activate naïve T cells [10]. Finally, effector T cells move to the B cell follicles of germinal centers (GCs) and secrete cytokines capable of promoting IgA class-switch recombination [11]. In mucosal immune effector sites such as the lamina propria of the gut, the upper respiratory tract, and the female reproductive tract, IgA+ plasma cells terminally differentiate to release secretory IgA (SIgA), the most important immune effector molecule in the mucosa. SIgA is transported across mucosal epithelial cells via a polymeric Ig receptor (pIgR) (Fig. 1) [12]. SIgA is a major immune effector at mucosal surfaces that acts via three mechanisms: antigen excretion, immune exclusion, and intracellular antigen neutralization (Fig. 2) [13]. Antigen excretion by SIgA features the binding of SIgA to pathogen-derived antigens, thus inhibiting pathogen–epithelial cell contact. SIgA exerts immune exclusion by eliminating antigens via secretion of an IgA–antigen complex, and invading pathogens can also be eliminated by complex formation with IgA-joining (J) chain-pIgR. SIgA inhibits the binding of pathogens and/or pathogenic antigens to specific receptors by neutralizing and eventually removing the pathogenic antigens.
Mucosal Vaccines and Delivery Routes
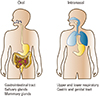
Mucosal vaccination can induce antigen-specific humoral and cell-mediated immune responses in both the systemic and mucosal compartments [14]. Additionally, such vaccination efficiently induces long-lasting B- and T-cell memory [15]. Importantly, the characteristics of mucosal immune response induction depend on the vaccine delivery route chosen (Fig. 3). For example, oral delivery (a traditional form of mucosal vaccination) can induce production of antigen-specific SIgA in the gastrointestinal tract, salivary glands, and mammary glands [14]. Currently, the licensed human live attenuated vaccines for rotavirus, poliovirus, Salmonella Typhi, and cholera are delivered orally (Table 3) [1416]. Intranasal vaccines such as FluMist, a live attenuated influenza virus vaccine, generate SIgA in the upper and lower respiratory, gastric, and genital tracts [17]. Upon sublingual vaccination, antigen-specific immune responses are induced in the gastrointestinal and the upper and lower respiratory tracts [14]. Although a few human mucosal vaccines are licensed, safety issues remain; the current vaccines are live attenuated or non-living whole-cell vaccines (Table 3). Subunit vaccines lacking entire pathogens are considered to be safer next-generation vaccines. However, several issues must be addressed when developing subunit mucosal vaccines, including poor immunogenicity, degradation of vaccine materials in the harsh mucosal environment, delivery of vaccine materials to mucosal immune inductive tissue, and modulation of the mucosal immune environment such that oral tolerance does not develop.
Mucosal Vaccine Adjuvants
The mucosa is continuously exposed to various antigens and microbiota and tightly regulates the influx of luminal antigens. Therefore, special delivery systems are required for development of successful mucosal vaccines [18]. M cells are the ideal targets of mucosal vaccine materials. Not only are the cells localized to the FAE of mucosal immune inductive sites, but many APCs are located nearby and/or under pockets of M cells. Although antigen uptake by M cells was previously thought to be non-specific, many recent studies have shown that a specific antigen delivery mechanism is involved [19]. GP2, a protein expressed specifically by M cells, drives transcytosis of FimH+ bacteria into such cells. Therefore, M cell-specific markers can be utilized for antigen delivery to mucosal immune inductive sites [2021222324252627282930313233343536373839] (Table 4). For example, an M cell-specific antibody, NKM 16-2-4, recognizes the α(1,2)-fucose-containing carbohydrate moiety of M cells and can be used to enhance delivery of an associated antigen [33]. Additionally, an M cell-targeting ligand, Co1, also targets antigens to M cells by interacting with the complement 5a receptor, inducing an antigen-specific immune response [2340]. Thus, M cell–targeting of vaccine materials will play a pivotal role in successful mucosal vaccination.
In the tolerogenic mucosal environment, adjuvants with immunostimulatory capacities enhance immune induction (Table 5) [37414243444546474849]. When TLR agonists such as Pam3CSK4, poly(I:C), MPL, or CpG-ODN were given either nasally or orally, in combination with vaccine materials, both systemic and mucosal antigen-specific immune responses were enhanced [3]. In addition, some immunostimulatory adjuvants improve the quality of the immune response. Cholera toxin (CT) is an effective mucosal vaccine adjuvant because it interacts with the GM1 ganglioside. However, the use of CT in this context raises a safety concern. Thus, CTA1-DD, which contains a mutant GM1 ganglioside-targeting A subunit of CT and the D-fragment of Staphylococcus aureus protein A to activate follicular dendritic cells (FDCs) closely associated with GCs, has been developed. CTA1-DD effectively promotes the induction of high-affinity B-cell clones and long-lived memory B cells and plasma cells [50]. Another mucosal vaccine adjuvant is the oil-in water emulsion MF59, which is currently licensed for human use. Although the mechanism of action remains unclear, MF59 not only enhances recruitment of innate immune cells via release of ATP and antigen uptake, but it also increases the adjunctive capacities of B cells by enhancing GC actions via activation of follicular helper T cells [51]. Finally, cathelicidin LL-37 is an immunostimulatory adjuvant that targets antigens to M cells. LL-37 increases antigen delivery to such cells and activates FDCs by interacting with the formyl peptide receptor 2 [24]. This enhances the induction of antigen-specific immune responses in both the systemic and mucosal compartments.
Conclusion
Recently, the need for mucosal vaccines has become recognized. Such vaccines offer several advantages including safety, convenience of vaccination, economical production, induction of mucosal immune responses, and enhanced memory B- and T-cell induction. However, several hurdles must be overcome in the development of practical subunit mucosal vaccines, including poor immunogenicity, degradation of vaccine materials in a harsh mucosal environment, delivery of vaccine materials to mucosal immune inductive tissue, and modulation of the mucosal immune environment to ensure that oral tolerance does not develop. These obstacles will be overcome by developing effective mucosal adjuvants that target M cells and are immunostimulatory.





 XML Download
XML Download