

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
During early mammalian development embryos possess the capacity to produce complete female and male reproductive tracts. The Müllerian ducts will become the upper third of the vagina, the cervix, uterus, Fallopian tubes and the outer lining of the ovaries. The Wolffian ducts become the seminal vesicles, vasa deferens and epididymides. These ductal systems grow adjacent to but independent of one another. Once the genetic sex of the embryo becomes known specific changes begin to occur. For example, at about 10 weeks gestation in human the undifferentiated gonads become either testes, under the control of the SRY gene [1] or ovaries. As gonads begin to differentiate one of the reproductive tract primordia is destroyed and the other proliferates and differentiates. In females, the lack of testosterone is sufficient to allow the Wolffian ducts to atrophy while in males the ablation of the Müllerian duct is more complex. The French anatomist Dr. Alfred Jost shed some very important light on the process in males over 65 years ago [2,3]. At that time it was believed that the key to normal prenatal sexual development was whether testes were present. The testosterone they secrete would both stimulate the Wolffian ducts and destroy the Müllerian ducts. Professor Jost's experiments proved that testosterone alone was insufficient to the task because the gonadectomized embryos he treated with testosterone crystals had retained uteri at birth. He proposed that a "Müllerian inhibiting substance (MIS)" now also called anti-Müllerian hormone (AMH), is secreted from the newly formed testes and that it is the MIS/AMH which causes the regression of the Müllerian ducts. It was later shown that regression is due mainly to apoptosis, autophagocytosis, disruption of basement membranes, and epithelial mesenchymal transformation of cells followed by migration of some of the cells toward the mesonephros [4,5,6,7,8,9].
Müllerian inhibiting substance/anti-Müllerian hormone protein
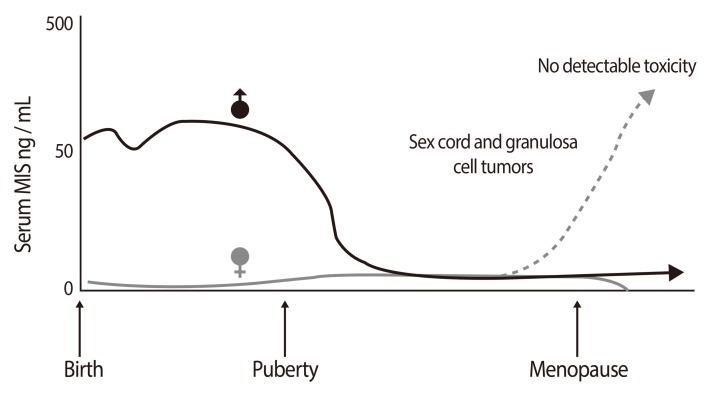
Over two decades after professor Jost's observation the source of MIS/AMH in males was identified as the Sertoli cells [10,11] in rat [12], bovine [13,14] and human testis [15,16]. In addition, it was learned that MIS/AMH production is sexually dimorphic. MIS/AMH gene transcription is regulated in males by a number of factors including SF1, SOX 9 and WT-1 in utero but after birth androgens have been implicated in suppression of MIS/AMH expression [17,18,19,20,21,22,23] while there are data showing MIS/AMH can inhibit androgen synthesis as well [24,25,26]. MIS/AMH is expressed in significant amounts only by males during fetal life but postnatally, both granulosa cells in the female and Sertoli cells in the male secrete MIS/AMH. In males serum MIS/AMH levels remain high until puberty, dropping to basal levels, thereafter. Although MIS/AMH mRNA is detectable in ovaries shortly after birth [27] MIS/AMH is undetectable in serum until puberty where its levels reach those of adult males (Fig. 1) [28]. The detection of MIS/AMH in both sexes after birth [29,30,31] suggested multifunctional roles for MIS/AMH. In females, MIS/AMH blocks meiosis II [32], inhibits ovarian granulosa cell division and progesterone production [33], and modulates follicular development [34]. Furthermore, MIS/AMH levels in follicular fluid are inversely correlated with granulosa cell proliferative index [35,36]. In males, MIS/AMH affects Leydig cell development and inhibits transcription of the steroidogenic enzyme, CYP17 [24,25,37,38]. Its stage-specific expression in seminiferous tubules implies a role in spermatogenesis [39]. Recent findings showing MIS/AMH and its receptor expression in pituitary and motor neurons indicate even more widespread non-Müllerian roles for MIS/AMH [40,41]. In fact, the prostate [42,43], mammary gland [44,45] and some of their cancers [45,46] as well as ocular melanoma [47] are possible targets for MIS/AMH. The role MIS/AMH plays in these tissues is at present unknown.
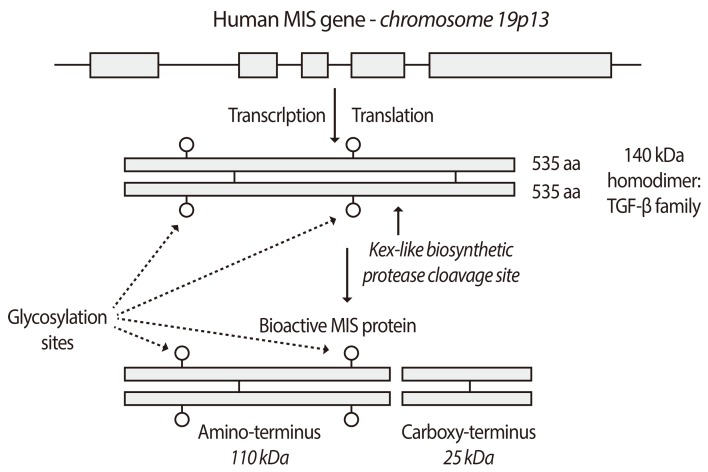
MIS/AMH was purified from testicular secretions in vitro [13,48,49] or protein extracts [13,50,51,52,53,54,55,56] by conventional biochemical techniques [13,48,49,50,51,52,53,54,55,56] and compositional analyses revealed it to be a 140 kDa glycoprotein disulfide linked homo-dimer of approximately 15% carbohydrate by weight. MIS/AMH is cleaved into two fragments, most likely during the biosynthetic process before secretion by kex-like prohormone convertases.
Analyses of bovine cDNA and human genomic sequences revealed MIS/AMH to be weakly related to the transforming beta family of proteins with the most striking homology (28%) residing in the carboxy-terminal domain [55,56]. The 2.8 kb human gene contains five exons and four introns and is located on the short arm of chromosome 19 [57]. The deduced protein sequence of the MIS/AMH monomer contains a 25 amino acid secretion specific signal peptide and a monomeric protein of 535 amino acids that, when glycosylated at two glycosylation sites, has a molecular weight of 70 kDa (Fig. 2) [28]. Lysis at a protease cleavage motif at amino acid residue 427 explains the origin of the major MIS/AMH fragments (55 kDa amino-terminus and 12.5 kDa carboxy-terminus) appearing on reduced polyacrylamide gels of the purified protein. A weaker motif at residue 229 is also present but its role in MIS/AMH action is unclear. In the native molecule it is the carboxy-terminal disulfide linked homodimer (25 kDa), which possesses the biological activity [58,59,60]. The amino-terminal domain may contribute to proper protein folding and assembly during synthesis and/or it may increase the serum half-life of the carboxy-terminus and enhance bioactivity [59]. The carboxy-terminal sequence of MIS/AMH is extremely highly conserved, a fact that explains why MIS/AMH from many different species are all active in the rat in vitro bioassay used for MIS/AMH purification. The human gene was transfected into Chinese hamster ovary (CHO) cells and its conditioned medium is used as a source of recombinant human MIS/AMH [61,62] for purification of the MIS/AMH needed for all of our studies. The MIS/AMH as purified from serum free conditioned media is proteolytically processed by endogenous CHO enzymes, it is free of contaminants from bovine serum, hamster cells, and the mouse monoclonal antibody used to purify it [62], and significant amounts of endotoxins [45].
Müllerian inhibiting substance/anti-Müllerian hormone receptors
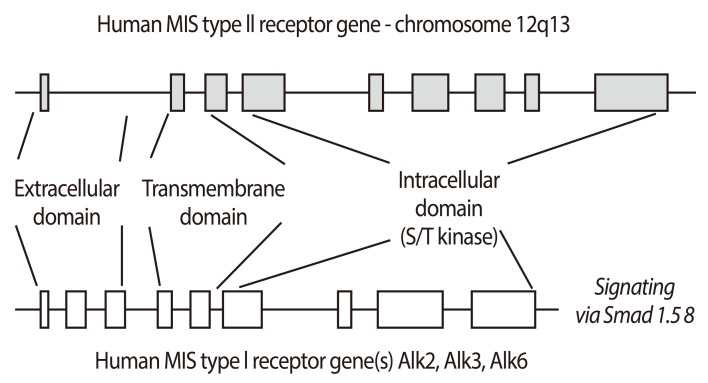
MIS/AMH biological activity requires interaction with two similar but distinct receptors termed type I and II. These single membrane spanning, serine-threonine kinases cross phosphorylate upon MIS/AMH binding and initiate intracellular cascades that lead to the control of cell cycle regulating proteins and altered transcription of a number of genes, depending upon the target tissues. The type II receptor binds the ligand and the type I heteromer is the signaling receptor (Fig. 3) [28]. The MIS/AMH type II receptor was cloned in the laboratories of Baarends and Themmen, Josso and Cate, and Behringer, as well as our own laboratory [63,64,65,66]. The human gene is located on chromosome 12q13 [67] and has 11 exons and 10 introns.
The MIS/AMH type II receptor is expressed in Müllerian duct mesenchyme around the adjacent ductal epithelium, consistent with the fact that the fetal mesenchyme directs Müllerian duct regression [63,64,65]. The receptor is also found in fetal and adult Leydig and granulosa cells [38,39,68,69] where it regulates testosterone [24,38,39,68], estradiol and progesterone synthesis [69]. The MIS/AMH type II receptor is made by rodent uteri [66,70], human endometrium [70], breast and prostate tissues [42,44] and, surprisingly, in motor neurons in the mouse brain [41]. It is not yet understood what MIS/AMH is doing to these tissues but MIS/AMH inhibits the proliferation of tumor cell lines derived from them supporting our hypothesis that MIS/AMH may be a useful adjuvant agent in the treatment of these diseases.
Persistent Müllerian duct syndrome results from loss of function mutations in the MIS/AMH type II receptor and/or the MIS/AMH molecule itself [71]. A common mutation in the MIS/AMH gene deletes the carboxy-terminal domain and is therefore, it fails to regress the Müllerian duct. Heterozygotes of either mutation type, i.e. receptor or ligand have normal phenotypes; affected individuals are either homozygous for a given mutation or compound heterozygotes. Recently, a new intron mutation associated with Persistent Müllerian duct syndrome has been identified, as well [72].
Serum Müllerian inhibiting substance/anti-Mü hormone clinical utility
The discovery that males and females both express MIS/AMH postnatally prompted numerous studies which revealed the usefulness of measuring MIS/AMH in serum in a variety of clinical situations including ambiguous genitalia, undescended testes, delayed puberty, sex cord and granulosa cell tumors. Fortunately, three very useful human MIS/AMH-specific and highly sensitive immunoassays were developed concurrently by the Donahoe, Josso and Hutson laboratories [29,30,31] for this purpose and several are commercially available thus, impacting the practice of pediatric endocrinology, pediatric surgery and gynecologic oncology [29,30,31].
Serum MIS/AMH, is a strong predictor of the presence of testicular tissue when testes are not palpable [79]. It also is a useful indicator of testis function in these cases and in cases of sexual ambiguity. Normal MIS/AMH levels are consistent with normal testes, undetectable MIS/AMH reflects absent testes whereas, low MIS/AMH is often due to gonadal dysgenesis. Serum MIS/AMH data can be more useful than hCG stimulation tests to evaluate gonadal status.
Serum MIS/AMH correlates with ovarian reserve and it is an indicator of response to in vitro fertilization protocols [36,80,81,82,83,84,85]. The vast majority of sex cord and granulosa cell tumors [86,87] secrete MIS/AMH in proportion to tumor burden [88] and changes in serum MIS/AMH reflect recurrences and/or response to therapy. These tumors may express the MIS/AMH receptor [89] but the tumors are not responsive to the MIS/AMH they produce. This fact implies a loss of function downstream from the receptor itself, therefore, these tumors are not considered targets for MIS/AMH therapy. Serum concentrations of MIS/AMH above the normal for age is consistent with the presence of either one of these tumors; rising serum MIS/AMH reflects tumor growth and decreases show response to therapy [86]. MIS/AMH levels can reach thousands fold the normal range without any adverse reactions that could be attributed to the MIS/AMH molecule. It is for this reason that we predict that MIS/AMH administered to cancer patients as a may have a very favorable toxicity profile (see below). Whether recombinant human MIS/AMH is toxic will be the topic of detailed toxicology studies planned for the future when clinical grade MIS/AMH is available.
Müllerian inhibiting substance/anti-Müllerian hormone and cancer therapy
Much of the research effort has focused the molecular mechanism of action of MIS/AMH [90,91,92,93]. A major goal of this work is to translate findings into new therapies or diagnostic tools for the management of pediatric diseases requiring medical intervention and/or surgical reconstruction. In addition to the clinical uses of MIS/AMH measurements listed above, an unexpected consequence of the knowledge gained form basic MIS/AMH research is the use of recombinant human MIS/AMH as a novel, naturally occurring, potentially nontoxic, cancer drug administered to patients with MIS/AMH receptor expressing malignancies.
As it became clear that MIS/AMH inhibits cell division and/or induces apoptosis in a highly specific manner, it was suggested by a colleague of ours, Professor Robert Scully of the Massachusetts General Hospital Pathology Department and an expert in ovarian cancer, that MIS/AMH might be an inhibitor of cancers of Müllerian duct origin. His observation that epithelial ovarian cancers recapitulate embryonic Müllerian histology [94], lead to the logical question of whether these lesions could be MIS/AMH targets. If so, administration of exogenous MIS/AMH could be an effective, highly specific reagent to include in existing treatments for epithelial ovarian cancer [95,96,97,98,99]. Results of more recent experiments, however, identify cervical and endometrial cancers as other potential targets [70,100], as well as several non-Müllerian cancers, including breast [43,45,46], prostate [43] and ocular melanoma [47] (see below). Our original hypothesis, therefore, is extended to include any tumor that expresses a functional MIS/AMH type II receptor. After examining a considerable body of pre-clinical experimental results we conclude that MIS/AMH may indeed be a useful cancer drug and we propose that phase I trials be initiated as soon as sufficient clinical grade material is available.
The concept that a naturally occurring growth inhibitor such as MIS/AMH could be an effective adjuvant treatment for cancer is attractive because MIS/AMH type II receptor expression is restricted to so few tissues thus minimizing the potential for adverse side effects. For the same reason, MIS/AMH may be used as a delivery system for more toxic drugs, again limiting exposure to non-target tissues. Our choice to focus initially on certain of the ovarian cancers arises from the fact that they have, when compared with other potential MIS/AMH targets, the worst prognosis and might benefit significantly from this novel biological reagent, thus offering a new approach to augment the efforts of others to find useful new strategies to employ.
1. Müllerian inhibiting substance/anti-Müllerian hormone and ovarian cancer
Validating MIS/AMH as an anti-cancer drug particularly for ovarian cancers has been the focus of our laboratory since we first hypothesized that any cancer of Müllerian origin could be a target for MIS/AMH treatment [95,96]. The notion that MIS/AMH could be used to treat ovarian cancer is predicted by the fact that the histology of the embryonic Müllerian ducts is somewhat reproduced in the common ovarian adenocarcinomas that can arise from the outer ovarian coelomic epithelium.
Epithelial ovarian cancer affects nearly 25,000 North American women each year and is the fifth most common malignancy in women with a five-year mortality of over 70% [101]. The mortality rate is significantly lower in stage Ia or Ib disease, but early peritoneal seeding and metastatic spread accounts for the fact that less than 25% of women are diagnosed with early stage disease. Combined surgery and cytotoxic therapy produce favorable clinical responses in 50% to 80% of patients but, unfortunately, the majority of patients relapse, therefore, it is timely to search for novel therapies for ovarian cancer [99,102].
Our hypothesis was supported by results of initial in vitro studies using human ovarian cancer cell lines [96,103,104,105] or tissues [106] followed by a series of ex-vivo experiments with human cancer cell lines [98]. Studies were also conducted on ovarian cancer cells in ascites specimens harvested collected from patients with new or recurrent disease [97]. Importantly, over 50% of the stage III ovarian patients had cells that bound recombinant human MIS/AMH, expressed the MIS/AMH type II receptor, and were growth inhibited by MIS/AMH [97]. Two other more recent studies corroborate and extend these findings on the incidence of receptor expression in benign and malignant cancers of Müllerian origin [107,108]. Therefore, the incidence of MIS/AMH responsive cases is sufficiently high to warrant further clinical investigation. The observations on fresh surgical specimens and ascitic fluid, however, are perhaps the most significant to date as they reflect what is true for patients and not cell lines. Receptor expression is frequent in a random sample of ovarian cancer cases and the receptor appears to be functional as MIS/AMH inhibits cancer cell proliferation in vitro [97].
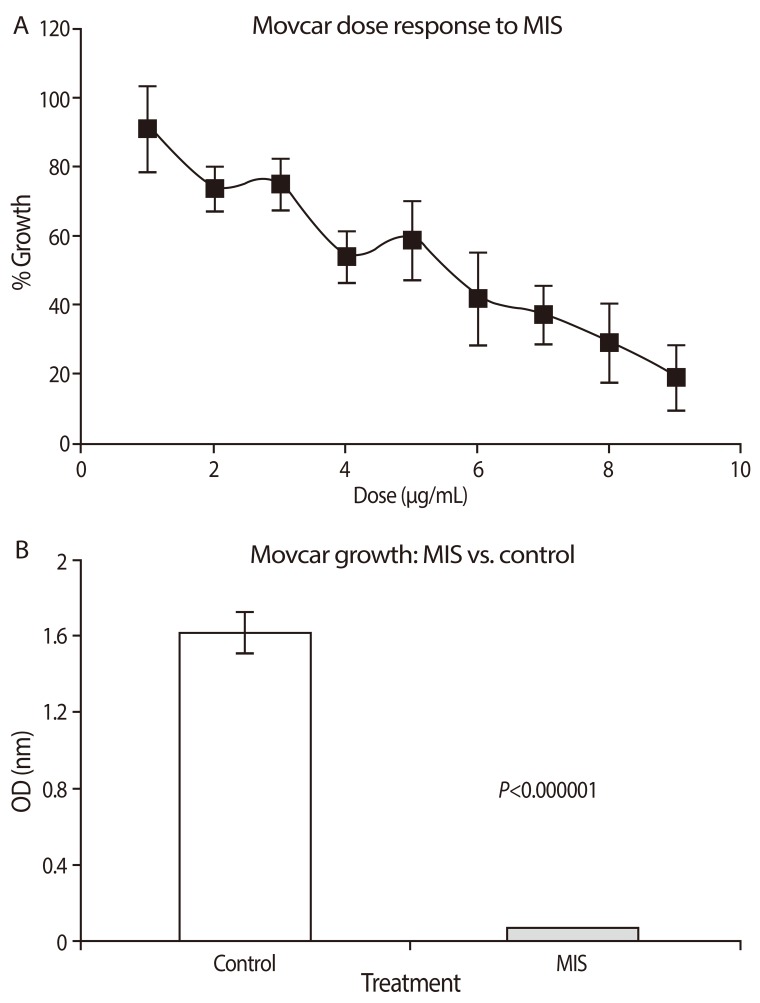
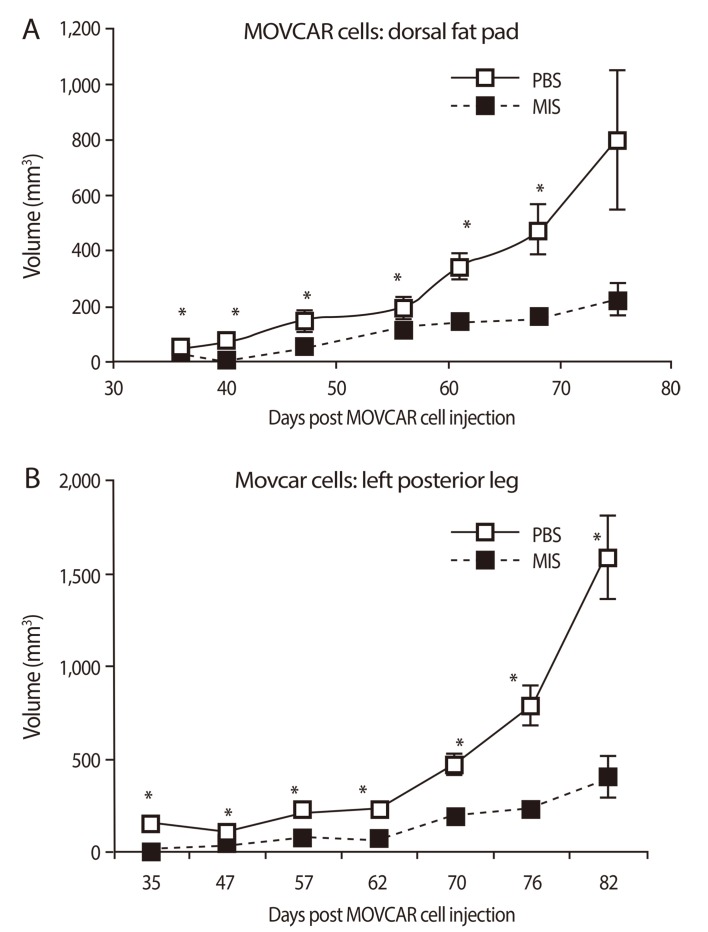
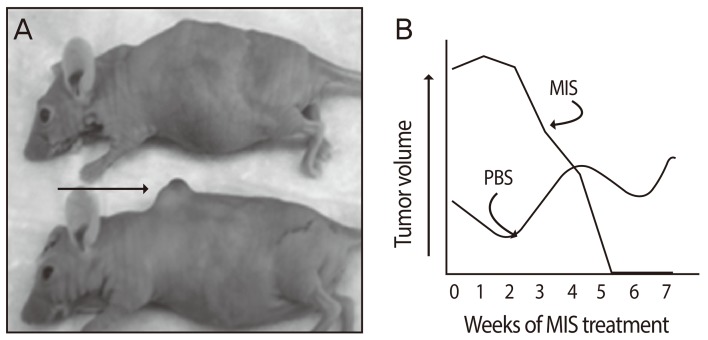
The pre-clinical studies of MIS/AMH in ovarian cancer were advanced greatly by the development of a new mouse model by Dr. Connolly and colleagues at the Fox Chase Cancer Center [109]. These tumors appear very similar to the phenotype of the most common human ovarian cancer, serous cystadenocarcinoma with a relatively large percentage (50%) of mice developing cancers within a few months of birth. Cell lines from these tumors, called MOVCAR cells, express the MIS/AMH type II receptor mRNA, they grew rapidly in culture, and they respond to MIS/AMH in vitro in a dose dependent manner with up to 95% growth inhibition (Fig. 4) [110]. To study the effects of MIS/AMH in vivo, nude mice were injected in the leg or into the dorsal fat pad with MOVCAR tumor cells at six weeks of age. Tumors appeared in a dose dependent manner in both implantation sites. When 10,000 cells were injected tumors appeared in approximately ten weeks. By comparison, 100,000 cells produced tumors in eight weeks, and tumors were visible in nine weeks after injection of 3×106 cells, and 12×106 cells produced tumors in approximately four weeks. Intraperitoneal injection of the animals with MIS/AMH lengthened the time to tumor appearance and tumor volume significantly smaller (Fig. 5). No apparent toxicity was detected and considering the fact that this duration of MIS/AMH exposure in the mouse is roughly equivalent to about 7 years of continuous cancer treatment in human, MIS/AMH may be relatively safe. In a preliminary experiment, a MOVCAR tumor was allowed to grow for over 20 weeks in the fat-pad before MIS/AMH treatment. The tumor completed regressed after 5 weeks of treatment days of treatment (Fig. 6) [111]. The buffer control treated animal's tumor continued to grow.
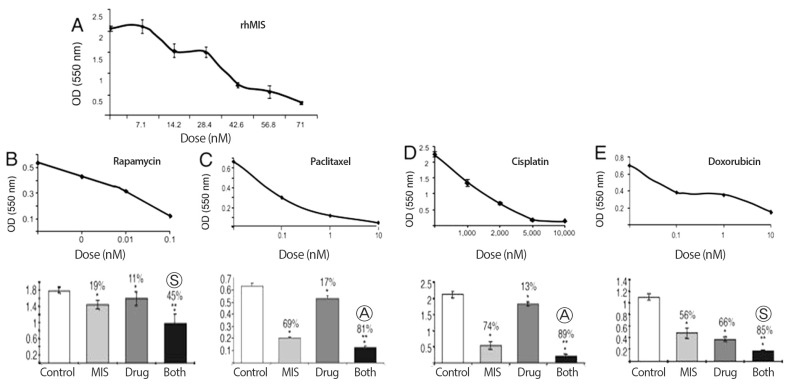
After phase I trials are completed MIS/AMH will most likely be administered to patients in simultaneously with other drugs. It was important, therefore, to study potential interactions between MIS/AMH and commonly used cytotoxic agents. Mechanism of action studies thus far show that MIS/AMH downstream signal transduction pathways include Smads, cyclin dependent kinase inhibitors, and cytokine inducible pathways. MIS/AMH also increased expression of p16, an inhibitor of cell cycle progression, the pocket proteins p107, and p130 as well as the apoptosis associated-E2F1 protein [105]. Another study reported last year has identified a number of other genes that are up and down regulated by MIS/AMH in an ovarian cancer cell line [112]. These results suggest MIS/AMH mechanisms of action very different from those of targeted by most chemotherapeutic agents. If MIS/AMH and drugs can function in combination it may be possible to decrease the dose needed for either agent alone, potentially resulting in decreased toxicity. In fact, preliminary studies with epithelial ovarian cancers in vitro demonstrated that combination therapy with MIS/AMH may indeed provide a means of reducing toxicity while maintaining or enhancing clinical effectiveness (Fig. 7) [113]. Using MOVCAR cells from the murine model dose responses to rapamycin, and three ovarian cancer chemotherapeutic drugs paclitaxel, cisplatin, and doxorubicin were established and the effect of added recombinant human MIS/AMH determined. These studied showed that rapamycin and MIS/AMH were synergistic; MIS/AMH and paclitaxel were additive; MIS/AMH and cisplatin were additive; and the combination of doxorubicin and MIS/AMH was synergistic.
The ultimate translation of MIS/AMH as a therapeutic, from studies on cancers in vitro, to preclinical trials in animals, and eventually to phase I clinical trials, means sufficient quantities of clinical grade recombinant human MIS/AMH need to be produced. Accordingly, our laboratory has spent considerable effort optimizing purification protocols from mammalian cells and is exploring a number of alternative sources for enhanced production of MIS/AMH. In addition, we have developed MIS/AMH-specific assays to detect the protein and measure its bioactivity with a high degree of precision.
2. Müllerian inhibiting substance/anti-Müllerian hormone and other Müllerian tissues and cancers
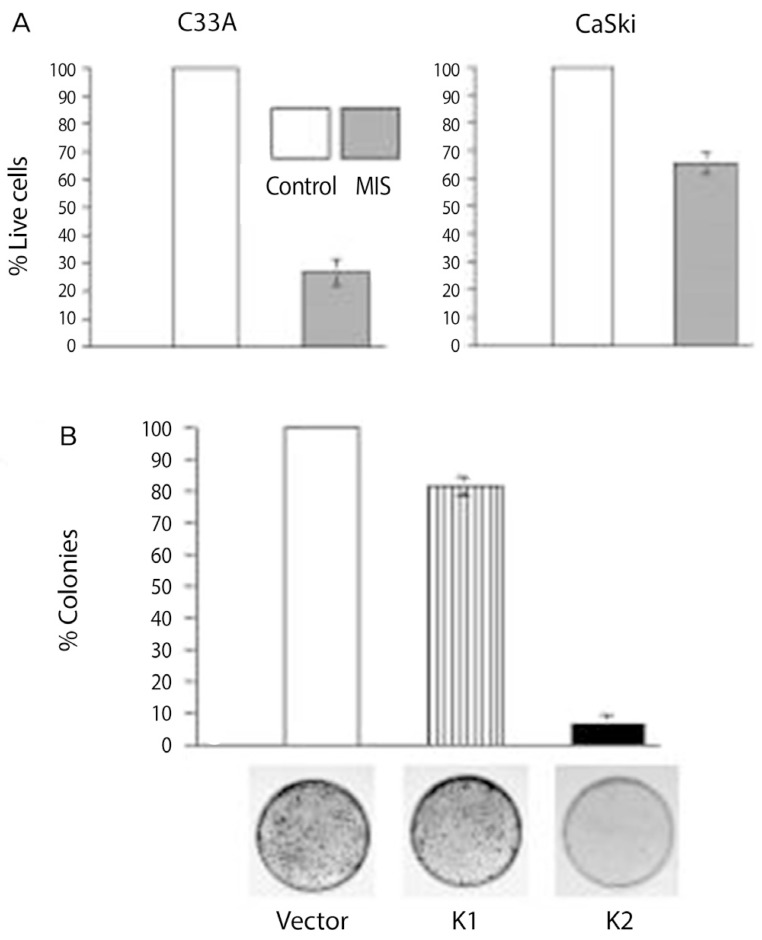
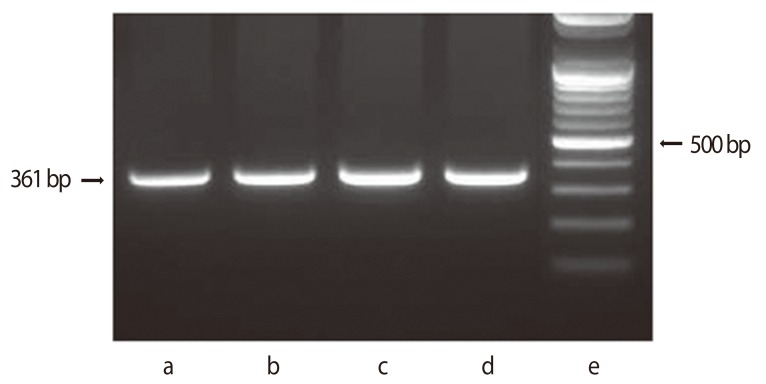
Normal cervix and cervical tumors express the MIS/AMH type II receptor protein by immunohistochemistry and mRNA by RT-PCR (Fig. 8) [100]. Human cervical cancer cell lines respond to MIS/AMH. Three human cervical carcinoma cell lines, CaSki, SiHa, and C33A were all growth inhibited in vitro by MIS/AMH added to the growth media or MIS/AMH produced by the cells themselves after transient transfection with a transcript encoding bioactive MIS/AMH (Fig. 9) [114]. As is true of ovarian cancer cell lines MIS/AMH upregulated p16, p130, p107, E2F1 in the C33A cell line. Interestingly, overexpression of p16, p130, p107, or E2F1 inhibited cell growth compared to vectors alone [100]. These findings in cervical cancer further broaden the targets for MIS/AMH.
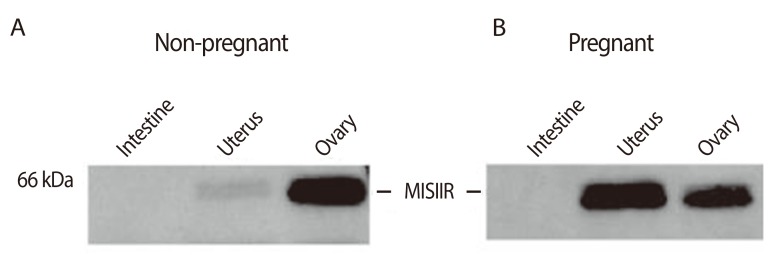
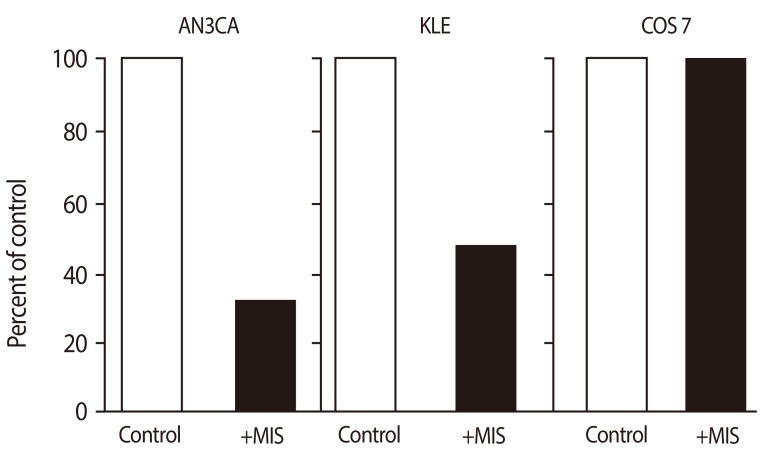
In addition to the cervix, Fallopian tubes, and the ovarian lining, the uterus is also a Müllerian structure. The MIS/AMH type II receptor mRNA is expressed in the Müllerian ducts of the female rat embryo between days 13 and 19 of development and in the uterine mesenchyme during the first postnatal days. The MIS/AMH type II receptor protein was also detected in a human endometrial cancer cell line AN3CA [70] and normal and pregnant adult rat uterus by western analysis [70] but not in small intestine (Fig. 10) [70]. In the human, MIS/AMH type II receptor mRNA was detected in myometrium and endometrium of a post-menopausal uterus and the protein observed by western analysis in endometrial tumors. Two cancer cell lines which express the receptor mRNA and protein were inhibited by MIS/AMH as well (Fig. 11) [70]. AN3CA, an endometrial carcinoma metastasis, showed significant growth inhibition (57% to 67% inhibition relative to control, 0%). KLE, a human cell line derived from a poorly differentiated endometrial cancer, also demonstrated significant growth inhibition (44.9%) after treatment with MIS/AMH. The cervical and endometrial transfection data support the conclusion that the growth inhibition effects are due to the MIS/AMH molecule itself rather than a contaminant in the purified protein preparation. Unlike the ovarian and cervical cancer cell lines AN3CA cells do not express p16 or p21 but MIS/AMH does induce p107, p130 and E2F1 [70].
3. Müllerian inhibiting substance/anti-Müllerian hormone and non-Müllerian tissues and cancers
It is noteworthy that MIS/AMH type II receptor is expressed in locations other than Müllerian tissues and gonads. In addition to the ocular melanoma already mentioned, MIS/AMH type II receptor is expressed in motor neurons [41] and the normal rat breast epithelium at levels inversely proportional to the state of proliferation of the tissue [44]. In several estrogen receptor positive and negative human breast cancer cell lines MIS/AMH enhanced IκB dependent DNA binding of NFκB that resulted in induction of IEX-1 mRNA [44], an immediate early gene induced by radiation, interferon γ, or tumor necrosis factor alpha but not by TGFβ [45,116,117]. Apoptosis was enhanced in the mammary epithelium of MIS/AMH injected animals compared to control animals [44], therefore, breast cancer could well be another target for MIS/AMH therapy.
MIS/AMH treatment of cancer may also include the prostate [42,43]. The MIS/AMH type II receptor and two candidate MIS/AMH type I receptors [74,75,76,77,78] are expressed in a prostatic cancer cell line (LNCaP) as well as in human prostate tumors [42]. MIS/AMH blocks the growth of human prostate cancer cell lines and it induces IRF1, and interferon γ was that enhances the inhibiting effects of MIS/AMH in vitro and in vivo. Furthermore, because MIS/AMH suppresses testosterone production [13,37,38,39] it might indeed exert a double effect on prostatic cancer, a direct growth inhibition and an indirect effect by lowering testosterone.
Müllerian inhibiting substance/anti-Müllerian hormone and endometriosis
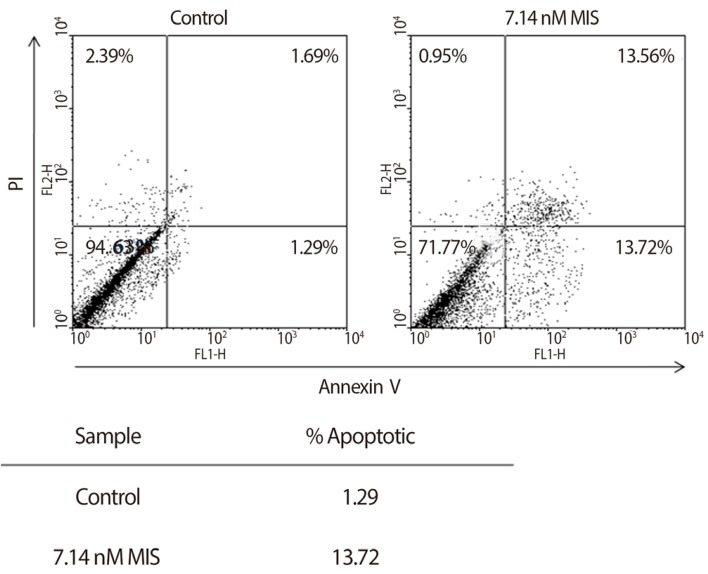
Endometriosis is a common benign disease of reproductive women, especially infertile women. Ectopic endometrial cells exhibit abnormal proliferative and apoptotic regulation in endometriosis. Recent reports show that MIS induces the cell cycle arrest and apoptosis of human endometrial stromal cells and endometriosis cell line in vitro. One study shows that the celluar apoptosis was increased by causing arrest at G1 phase and increasing cells at sub-G0G1 phase in endometrial stromal cells treated with MIS for 72 hours (Fig. 12) [118]. Another study shows that MIS induces autophagy through increased expression of Beclin-1 autophagy-promoting protein in endometriosis cell line [119]. These findings suggest that MIS play an important role in cellular apoptosis in endometriosis. Therefore, MIS has a potential as a therapeutic modalities in endometriosis.
Conclusion
The existing literature on MIS/AMH suggests that the recombinant human protein will have very little toxicity in vivo and that it should be tested against tumors expressing the MIS/AMH type II receptor, perhaps beginning with ovarian cancer as among receptor expressing tumors, since it has the most dire prognosis. The molecular tools to identify MIS/AMH receptor expressing ovarian and gynecologic tumors are already in hand, thus, it is possible to select patients for treatment. An MIS/AMH ELISA exists to follow administered doses of MIS/AMH, as well. Clinical trials await the production of sufficient supplies of recombinant human MIS/AMH for this purpose.
 XML Download
XML Download