

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Crohn's disease (CD) is a chronic inflammatory disease that typically affects the terminal ileum and/or colon. Despite extensive study, the pathophysiology of CD remains unclear; nonetheless, host genetic factors, immune system dysregulation, and environmental factors are known to be involved.1
2 Importantly, intestinal bacterial communities play a critical role in the pathogenesis of the intestinal inflammation that occurs in CD,3
4
5 and dysbiosis, a major change in the composition of intestinal microbiota, can contribute to increased immune stimulation and mucosal barrier dysfunction.4
We have reported that macrophages differentiated in vitro by macrophage colony-stimulating factor (M-CSF) and interferon-γ (IFN-γ) (Mγ-Mϕs) from plasma blood mononuclear cells show a similar phenotype to that of intestinal Mϕs.6
7 Mγ-Mϕs and lamina propria CD14+ macrophages have the potential to produce inflammatory cytokines, including tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6) and IL-23, and this phenotype resembles that of M1 macrophages. In contrast to Mγ-Mϕs, the characteristics of macrophages differentiated by M-CSF (M-Mϕs) alone are similar to those of M2 macrophages, including the ability to produce a large amount of IL-10. Using these in vitro differentiated macrophages, we recently found that a receptor for bile acid signaling modulates immune response in CD patients.8
Anti-Saccharomyces cerevisiae antibodies (ASCAs) are known to be directed against the component of the yeast, and that has been proposed as serological markers for CD diagnosis.9 Importantly, Candida albicans has also been shown to generate ASCAs.10 Fungi, including C. albicans, represent another major community of the human intestine microbiota, and recent research has highlighted the importance of the fungi residing in the gut and the interaction between these resident fungi and intestinal inflammation.11
12
13
14
15
16
17
Candida species were found to be significantly more abundant in the feces of IBD patients compared to healthy controls,11
13
14
15
16
17 and another study indicated that bacterial diversity was increased and fungal diversity reduced in pouchitis patients who maintained clinical remission with probiotics.12 β-(1,3)-Glucans represent 40% of the cell wall of the yeast and C. albicans. C-type lectin receptors, including Dectin-1 and Dectin-2, recognize fungi and are involved in host defense mechanisms by initiating inflammatory and adaptive immune responses.18
19 Dectin-1 recognizes β-(1,3)-glucans and induces innate immune responses,20 whereas Dectin-2 mainly recognizes α-mannose.21 The effect of Candida on murine colitis has also been investigated. For example, infection significantly delayed colonic healing in mice,22 and it was also reported that colonization by C. albicans is promoted by DSS colitis in mice and enhances inflammatory responses through galectin-3.23
Because most investigations of the interaction between fungi and host cells have been performed using murine models, it is not clear whether C. albicans influences inflammation in human intestinal macrophages in a manner that impacts CD pathogenesis. Here, we investigate the response of macrophages to curdlan (β-(1,3)-glucan), one of the cell wall components of commensal fungi. The aim of this study is to reveal the interaction between commensal fungi and host immune cells in IBD.
METHODS
1. Collection of Peripheral Blood Cells and Intestinal Mucosa from Patients with CD
A total of 15 mL of peripheral blood was obtained from patients with CD and healthy controls. Colonic mucosa samples were obtained from surgically resected specimens from patients with CD and UC patients for this study. For all samples from CD patients, mucosa samples with macroscopically moderate to severe inflammation were obtained. All studies were approved by the Ethics Committee of Keio University Hospital (20090253) and written informed consent was obtained from all patients.
2. Isolation of Peripheral Blood Monocytes and In Vitro M and Mγ Differentiation
CD14-positive (CD14+) monocytes were isolated from peripheral blood mononuclear cells (PBMCs) using CD14+ MACS (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's instructions. The percentage of monocytes isolated using this method was evaluated by flow cytometry and was routinely >98%. For in vitro M-Mφ differentiation, CD14+ monocytes were cultured with 50 ng/mL recombinant human M-CSF (R&D Systems, Minneapolis, MN, USA) for 6 days. For in vitro Mγ-Mφ differentiation, CD14 monocytes were cultured with 50 ng/mL recombinant human M-CSF (R&D Systems) and 100 ng/mL recombinant human IFN-γ for 6 days.
3. Preparation of Lamina Propria Mononuclear Cells
Lamina propria mononuclear cells (LPMCs) were isolated from intestinal specimens using modifications of a previously described technique.6 Briefly, dissected mucosa samples were incubated in calcium and magnesium-free HBSS (Sigma-Aldrich, St. Louis, MO, USA) containing 2.5% heat-inactivated FBS (BioSource International, Camarillo, CA, USA) and 1 mM DTT (Sigma-Aldrich) to remove mucus. The mucosa was then incubated twice in HBSS containing 1 mM EDTA (Sigma-Aldrich) for 45 minutes at 37℃. Tissues were collected and incubated in HBSS containing 1 mg/mL collagenase type 3 and 0.1 mg/mL DNase I (Worthington Biochemical, Lakewood, NJ, USA) for 60 minutes at 37℃. The fraction was pelleted and resuspended in 40% Percoll solution (Amersham Biosciences) and layered onto 60% Percoll before centrifugation at 2000 rpm for 20 minutes at room temperature. Viable LPMCs were recovered from the 40% to 60% layer interface.
4. Isolation of Lamina Propria Mφ
Lamina propria (LP) CD14+ Mφs were isolated from LPMCs using EasySep human CD14 (Stem Cell Technologies, Vancouver, Canada). The percentage of each subset of cells isolated using this method was evaluated by flow cytometry and was routinely >95%.
5. Commensal Fungal Heat-Inactivated Antigens
Commensal fungal strains of C. albicans were cultured on YPD agar. Fungi were harvested and washed twice with ice-cold PBS. Next, fungal suspensions were heated at 5℃ for 30 minutes, washed, resuspended in PBS, and stored at −80℃. Complete killing was confirmed by 72 hours incubation on solid medium at 37℃.
6. Quantitative Real-Time RT-PCR Analysis
Total RNA was extracted using an RNeasy Micro kit (Qiagen, Hilden, Germany), and cDNA was synthesized using a QuantiTect RT kit (Qiagen) according to the manufacturer's instructions. Quantitative real-time RT-PCR was performed using the TaqMan Universal PCR Master mix (Applied Biosystems, Foster City, CA, USA) and on-demand gene-specific primers for human dectin-1. The results were analyzed using DNA Engine Opticon 2 System (MJ Research, Waltham, MA, USA). Relative quantification was achieved by normalizing to the values of the β-actin gene.
RESULTS
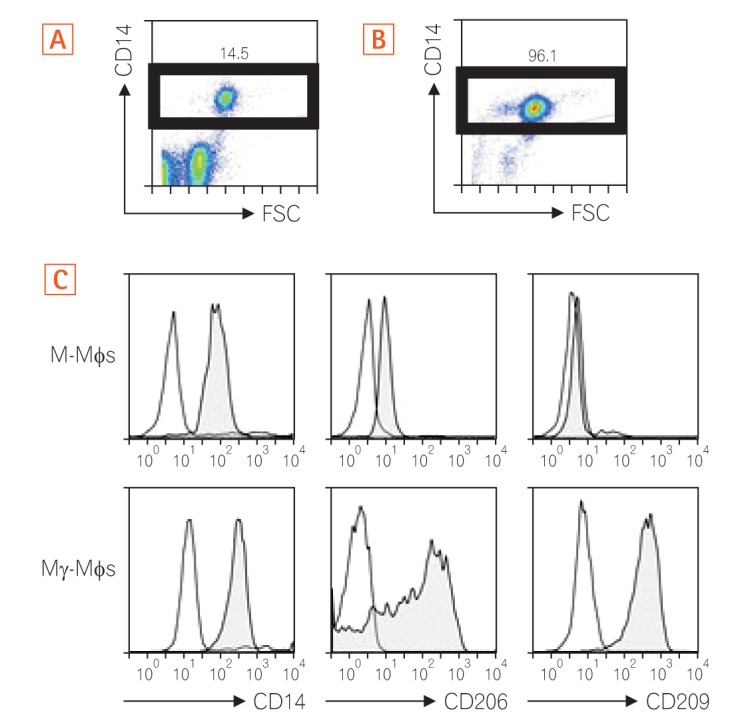
1. Differentiated Mγ-Mφs Express CD206 and CD209
As shown in Fig. 1A and B, we enriched CD14+ monocytes using CD14+ MACS. The enriched CD14+ monocytes were then differentiated into M-Mφs or Mγ-Mφs according to our previous report.24 We investigated the surface markers of these cells to characterize their surface phenotypes. The Mγ-Mφs expressed higher levels of the mannose receptor (CD206) and C-type lectin CD209 than the M-Mφs (Fig. 1C).
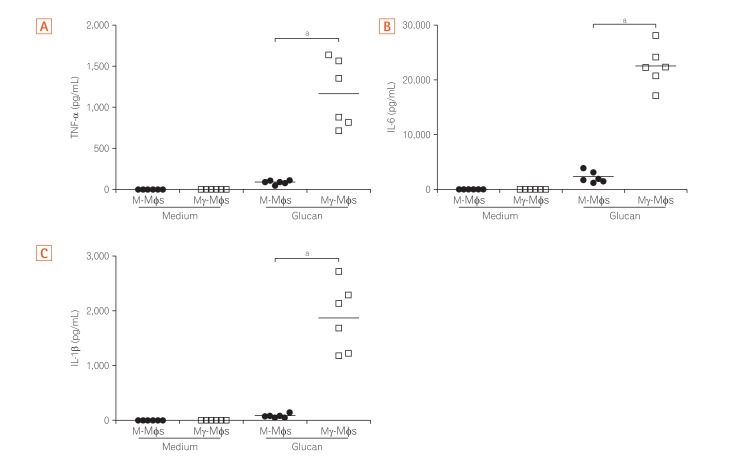
2. Mγ-Mφs Produce a Large Amount of Inflammatory Cytokines in Response to β-(1,3)-Glucan
The cytokine profiles of macrophages derived from the peripheral blood cells of healthy humans in response to β-(1,3)-glucan (purchased from Wako Pure Chemical; Product code #034-09901) were analyzed. Secretion of TNF-α, IL-6, IL-1β from both M-Mφs and Mγ-Mφs in the presence of glucan were higher than secretion in the absence of glucan (Fig. 2). The pro-inflammatory cytokine levels from Mγ-Mφs were significantly higher compared to those from M-Mφs (P<0.001), whereas secretion of IL-10, an anti-inflammatory cytokine, from Mγ-Mφs was lower than from M-Mφs in the presence of glucan (Supplementary Fig. 1). Interestingly, increase of TNF-α was in a concentration dependent manner in response to glucan (data not shown). These results suggest that the cytokine-producing potential of Mγ-Mφs is consistent with that of M1 macrophages, producing high amount of pro-inflammatory cytokines in response to fungal components.
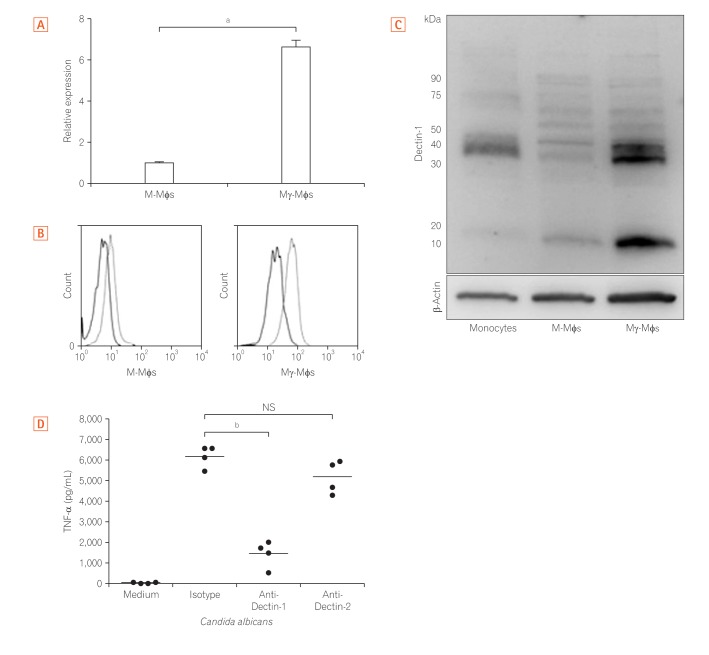
3. Glucan Induced Pro-inflammatory Cytokines in Mγ-Mφs via Dectin-1
Because the large amount of inflammatory cytokines produced by Mγ-Mφs was dependent on the concentration of β-(1,3)-glucan, we next assessed Dectin-1 expression. The mRNA levels of Dectin-1 in Mγ-Mφs were 6.7-fold higher than those in M-Mφs (P<0.01) (Fig. 3A). According to flow cytometry (Fig. 3B) and Western blotting (Fig. 3C), Dectin-1 expression was also markedly higher on Mγ-Mφs compared to M-Mφs. Previous reports have demonstrated that cytokines, including TNF-α, induction by β-glucans is dependent on Dectin-1 in bone marrow-derived dendritic cells25 and that Dectin-2 recognizes α-mannans and induces Th17 cell differentiation.26 TNF-α production by Mγ-Mφs was also examined and was markedly increased in the presence of heat-killed C. albicans. Furthermore, this higher production of TNF-α was largely reversed by an anti-Dectin-1 antibody but not an anti-Dectin-2 antibody (Fig. 3D). These results showed that Mγ-Mφs produced TNF-α through Dectin-1 not Dectin-2.
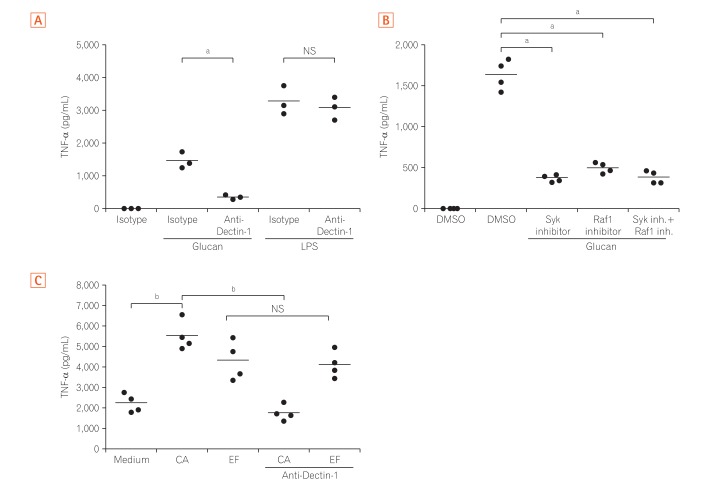
4. Dectin-1 Dependent Pathway Induces TNF-α Production in Mγ-Mφs and LP Macrophages from CD Patients
Next, TNF-α production by Mγ-Mφs, which were cultured in the presence of an antibody against Dectin-1, was analyzed to clarify whether the large amount of TNF-α produced by Mγ-Mφs occurs via interaction between glucan on the surface of various fungi and Dectin-1. The increased production of TNF-α by glucan-stimulated Mγ-Mφs was reversed by blocking Dectin-1, and this effect was not observed when Mγ-Mφs were co-cultured with lipopolysaccharide (LPS) (Fig. 4A). Although Gow et al.27 showed that spleen tyrosine kinase (Syk) inhibitors could not suppress TNF-α production in human PBMCs, TNF-α production by Mγ-Mφs was reversed by Syk or Raf-1 inhibitors (Fig. 4B). Taken together, TNF-α production by Mγ-Mφs occurs via Dectin-1-Syk/Raf-1 pathway, but not LPS dependent pathway.
Because we previously reported that Mγ-Mφs mimicked the phenotypes of intestinal macrophages in patients with CD,7 we next assessed cytokine production by LPMCs from patients with CD. TNF-α production by CD3–CD14+ LPMCs isolated from CD patients was high (2,076±323 pg/mL), even under un-stimulated conditions. TNF-α levels were also significantly higher under the condition of co-culture with C. albicans and Enterococcus faecalis compared to the unstimulated condition (P<0.01) (Fig. 4C). Although the anti-Dectin-1 antibody decreased TNF-α production by LPMCs when co-cultured with C. albicans, this effect was not observed after stimulation with E. faecalis. These results suggested that TNF-α production of LP macrophages induced by C. albicans in patients with CD was via Dectin-1 dependent pathway as Mγ-Mφs in vitro.
Because we previously reported that LP macrophages possess unique characteristics and influence pathogenesis of CD,7 we hypothesized that these TNF-α production pathways via Dectin-1 contribute to the pathogenesis of CD. We compared TNF-α and IL-6 production by LPMCs from patients with CD and UC using LPMCs isolated from surgical specimens. IL-6 production by LPMCs from patients with CD was significantly higher than in cells from patients with UC. In CD, IL-6 production by LPMCs stimulated with C. albicans was higher than by cells not stimulated with C. albicans. However, the difference was not significant (P=0.12). Although TNF-α production by LPMCs stimulated with C. albicans was higher for UC patients, this response was more than 10 times higher for CD patients (12,347±894 pg/mL) than UC patients (786±143 pg/mL) (Supplementary Fig. 2).
DISCUSSION
Although a number of studies investigating the role of microbiota,28
29
30
31 including commensal fungi,22
23 in intestinal inflammation have been conducted in models of experimental colitis, there are few studies on the contribution of commensal fungi to intestinal inflammation in patients with CD. Here, we demonstrate, for the first time, that C. albicans activated the production of TNF-α via Dectin-1, and not Dectin-2, in both in vitro differentiated macrophages and LP macrophages from patients with CD.
The role of glucan as a pro-inflammatory or anti-inflammatory stimulus is an ongoing debate. One study indicated that mice lacking Dectin-1 exhibited an increased susceptibility to chemically induced colitis, which was the result of altered responses to indigenous fungi. Furthermore, a polymorphism in the gene encoding Dectin-1 (CLEC7A) is strongly linked to a severe form of UC.32 These results suggest that the interaction between fungi and Dectin-1 expressed by immune cells influences health and protects against inflammatory diseases. The present studies indicate that glucan induced production of pro-inflammatory cytokines via dectin-1 on macrophages and our results are consisted with recent studies indicating that bacteria-derived glucan induced enteritis in murine models of IBD.22
23 These results may reflect that glucan of different origin-fungal versus bacterial- has different effects on enteritis.
Several studies also indicate that the composition of symbiotic fungi in the gut differs between CD patients and healthy controls. Importantly, the number of Candida genera in CD patients was increased compared to healthy controls.11
13
14
15
16
17 More recently, fungal dysbiosis was observed, with an increased Basidiomycota/Ascomycota ratio, a decreased proportion of S. cerevisiae and an increased proportion of C. albicans compared with healthy controls.17 Another recent study also indicated that Candida glabrata was overrepresented in CD and that S. cerevisiae and Filobasidium uniguttulatum were associated with non-inflamed mucosa in CD.15
Candida was also significantly more abundant in stool samples from pediatric patients with IBD, and it was also confirmed that pediatric IBD is associated with reduced diversity in both fungal and bacterial gut microbiota.11 These results suggest that fungi, including C. albicans, might play a role in pathogenesis of CD. However, these studies did not confirm whether commensal fungi interact with inflammatory cells within the intestinal lamina propria in humans.
In our study, we demonstrated that C. albicans affected the production of pro-inflammatory cytokines by CD3–CD14+ LPMCs from CD patients via Dectin-1. Saijo et al.25 demonstrated that Dectin-1 is not required for defense against C. albicans, suggesting that molecules other than β-1,3- and β-1,6-glucans are mainly involved in the induction of cytokines after infection of this fungus. Brown and Gordon33 discovered that Dectin-1 is expressed in each macrophage population. Dectin-1 induces cytokine production through the following 3 pathways: Toll-like receptor-Myd88-dependent production,20 Syk-dependent production,34
35
36 and Raf-1 dependent production.34 Furthermore, a recent study suggests that interactions between commensal fungi and Dectin-1 influence colitis.32 In our study, inhibition of Dectin-1, Syk, and Raf-1 failed to completely suppress TNF-α production, which suggested the existence of another pathway for TNF-α production.
In the murine model, colonization by C. albicans increases IL-17 and IL-23 production by gastric and oral tissues,37
38 indicating that Candida colonization might enhance inflammation by increasing the levels of these cytokines.13 Other recent studies indicate that this fungus exacerbates damage and delays healing of inflammatory lesions in animal models.22
23
C. albicans makes contact with intestinal epithelial cells and can also interact with immunologically competent cells in the gut. In the present study, we assessed pro-inflammatory cytokine production by LPMCs stimulated with heat-killed C. albicans in patients with UC and CD. Interestingly, TNF-α production was significantly higher in LP macrophages from CD patients compared to UC patients when the cells were stimulated with C. albicans. These results suggest that the responsiveness of LPMCs to the fungus differs between CD and UC, and thus, these antigens may differ in their contribution to disease.
This is the first study to show that LPMCs derived from CD patients stimulated with β-(1,3)-glucan produce a large amount of TNF-α compared with cells derived from UC patients. The findings indicate the possibility of utilizing C. albicans as an indicator of chronic inflammation in patients with CD. Although several therapies have been developed within the last decade, CD patients who are primary refractory or who have lost secondary responsiveness to medical treatment are still observed in the clinical setting. As recent report suggested,39 we also indicate that improvement of fungal dysbiosis and Dectin-1-Syk-Raf-1 signaling pathway may be a therapeutic target in CD.
 XML Download
XML Download