

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Stress can have various effects on the human biological systems. Stress is omnipresent in modern life and has great effect on cognitive and emotional functions [1]. Stress is defined as a state of disharmony or threatened homeostasis and results in various physiological and behavioral changes. The immobilization stress model is the most commonly employed models to mimic potent physical and psychological stress [2]. Moreover, the neuroanatomical regions mediating stress response, involving a variety of neurotransmitter systems, have been identified. Among all the brain regions, the PVN is a region that plays a central role in the regulation of the HPA axis during the stress [3,4]. It has been characterized that stress influences brain activity and promotes long-term changes in various neural systems [5]. For example, stress stimulates the release of various hormones, these increased blood glucose levels [6]. Stress leads to the activation of sympathetic nervous system (SNS) and hypothalamic-pituitary-adrenal (HPA) responses [7]. The SNS activation produces catecholamines that are known as major stress hormones, especially epinephrine and norepinephrine. Increased activity of the SNS plays a role in the regulation of glucose metabolism. The alterations in glucose and lipid metabolism induced by stress may cause the development of diabetes mellitus [8]. The main regulator of the HPA axis is a corticotrophin-releasing hormone, finally causing a glucocorticoid secretion by the adrenal cortex [9]. Glucocorticoid usually increases the gluconeogenesis activity, leading to an increase of the blood glucose level [10].
Sulfonylurea is widely used oral drugs for the treatment of type II diabetes mellitus. Many studies have also reported that the major blood glucose-lowering activity of sulfonylurea appears to be primarily through enhance β-cell responsiveness. Also, increase intracellular cyclic AMP; exert insulinotropic effects by closing ATP-dependent potassium channels [11,12]. Its action of blood glucose is suppressed glucagon secretion, increase peripheral insulin sensitivity without affecting insulin binding [13]. Skillman and Feldman, reported that the sulfonylurea potentiate the biologic effect of the insulin, increasing the deficient numbers of insulin receptors and stimulating insulin secretion on skeletal muscle, fat and liver [14].
The peripheral anti-diabetic actions of sulfonylurea have been well studied and characterized. Chari et al. [15] demonstrated that the activation of central lactate metabolism after i.c.v administration of lactate, lowers the glucose production in uncontrolled diabetes and diet induced insulin resistance. Park et al. [16] have recently demonstrated that both morphine and β-endorphin administered i.c.v. acutely increase the blood glucose level. Furthermore, Sim et al. [17] reported that the anti-diabetic action of repaglinide appears to be, at least, mediated via the brain and the spinal cord as revealed in both D-glucose-fed and STZ-treated models. However, the roles of sulfonylureas directly administered into the brain in the regulation of stress-induced blood glucose level have not been well characterized yet. Thus, the present study was designed to examine the effect of sulfonylurea administered supraspinally on the blood glucose level induced by immobilization stress in mice.
METHODS
These experiments were approved by the Hallym University Animal Care and Use Committee (Registration Number: Hallym 2009-05-01). All procedures were conducted in accordance with the 'Guide for Care and Use of Laboratory Animals' published by the National Institutes of Health and the ethical guidelines of the International Association for the Study of Pain.
Experimental animals
Male ICR mice (MJ Co., Seoul, Korea) weighing 20~25 g were used for all the experiments. Animals were housed 5 per cage in a room maintained at 22±0.5℃ with an alternating 12 h light-dark cycle. Food and water were available ad libitum. The animals were allowed to adapt to the laboratory for at least 2 h before testing and were only used once. Experiments were performed during the light phase of the cycle (10:00~17:00). The animals were fasted for 16 hours. The behavioral scores were verified by repeat trials, and the observers of pain-induced behavior were blind to drug treatments.
Intracerebroventricular injection
The intracerebroventricular (i.c.v.) administration followed the method described by Haley [18]. Each mouse was grasped firmly without anesthesia by the loose skin behind the head. The skin was pulled taut. A 30-guage needle attached to a 25 µl syringe was inserted perpendicularly through the skull into the brain and solution was injected. The injection site was 2 mm from either side of the midline on a line drawn through the anterior base of the ears. The i.c.v. injection volumes were 5 µl, and the injection sites were verified by injecting a similar volume of 1% methylene blue solution and determining the distribution of the injected dye in the ventricular space. The success rate for prior injections with this technique was over 95%.
Immobilization stress procedure
The mice were subjected to restraint stress as described in a previous study [19]. In brief, restraint was carried out by placing the mouse in a 50 ml corning tube, and adjusting it with an iron nail on the outside, which crossed in the caudal part of the animal. Adequate ventilation was provided by means of holes at the sides of the tubes. The mice were stressed by restraint for 30min. In the stress model there was a single exposure.
Measurement of blood glucose level
The blood glucose level was measured at 30, 60 and 120 min after the immobilization stress initiated. The blood was collected shortly as much as possible with a minimum volume (1 µl) from the tail-vein. The glucose level was measured using Accu-Chek Performa blood glucose monitoring system (glucometer) (Mannheim, Baden-Württemberg, Germany).
Corticosterone assay and blood sampling
The plasma corticosterone level was determined by the fluorometric determination method [20]. Four hundred microliters of blood were collected by puncturing the retro-orbital venous plexus. Plasma was separated by centrifugation and stored at 80℃ until assayed.
Insulin ELISA assay
In Mouse Insulin ELISA, biotin conjugated anti insulin, and standard or sample are incubated in monoclonal anti-insulin-coated wells. After horse radish peroxidase (HRP) conjugated streptavidin remaining in the wells are reacted with a substrate chromogen reagent and the reaction is stopped by addition of an acidic solution, and absorbance is measured spectrophotometrically at 450 nm.
Drugs
Tolazamide, glyburide, glipizide and glimepiride were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Tolazamide, glyburide, glipizide and glimepiride were prepared following steps: (A) 1 g of pioglitazone and rosiglitazone was dissolved in 0.5 ml of ethanol plus 0.5 ml of polyethylene glycol 400. (B) Separately, 100 mg of sodium carboxymethylcellulose was dissolved in 9 ml of distilled water. (C) Finally, Solution (A) and Solution (B) were vigorously mixed. This solution (PEC) excluding tolazamide, glyburide, glipizide and glimepiride were used as vehicle control. All drugs were prepared just before use. Blood glucose meter, lancing device and strips were purchased from Roche Diagnostics (Accu-Chek Performa, Germany).
Statistical analysis
Statistical analysis was carried out by student t test GraphPad Prism Version 4.0 for Windows (GraphPad Software, San Diego, CA, USA). p-values less than 0.05 were considered to indicate statistical significance. All values were expressed as the mean±S.E.M. In our study, we established the mean blood glucose value of the control group through many experiments under matching conditions. Selected mice of established blood glucose level were then used in replication experiments.
RESULTS
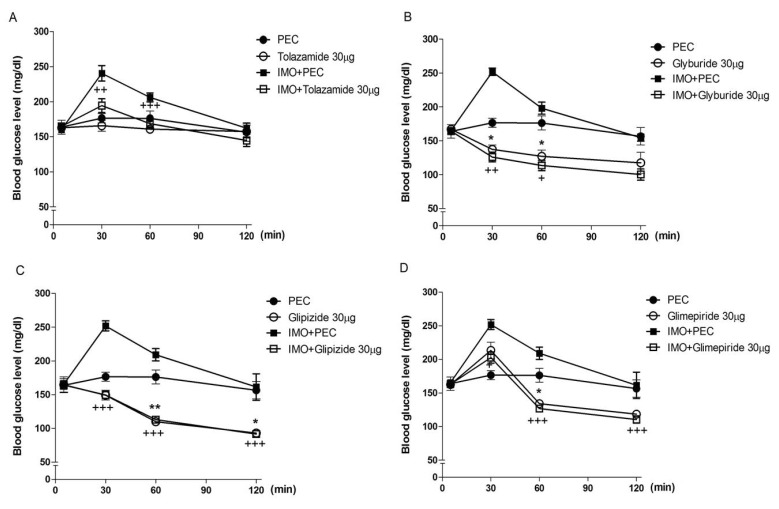
Effects of sulfonylureas administered intracerebroventricularly on the blood glucose level in immobilization stress model
Mice were pretreated i.c.v. with 30 µg of tolazamide, glyburide, glipizide or glimepiride for 10 min. Then mice were enforced into immobilization stress for 30 min and returned to the cage. The blood glucose level was measured at 30, 60 and 120 min after immobilization stress initiated. As shown in Fig. 1, tolazamide, glyburide, glipizide and glimepiride pretreated i.c.v. attenuated the elevation of the blood glucose level induced by immobilization stress, respectively.
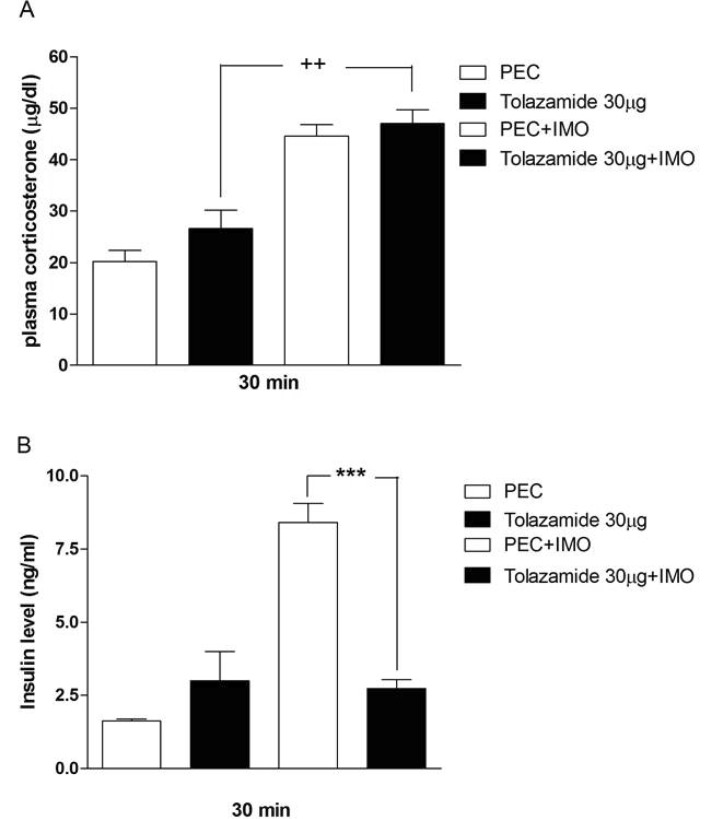
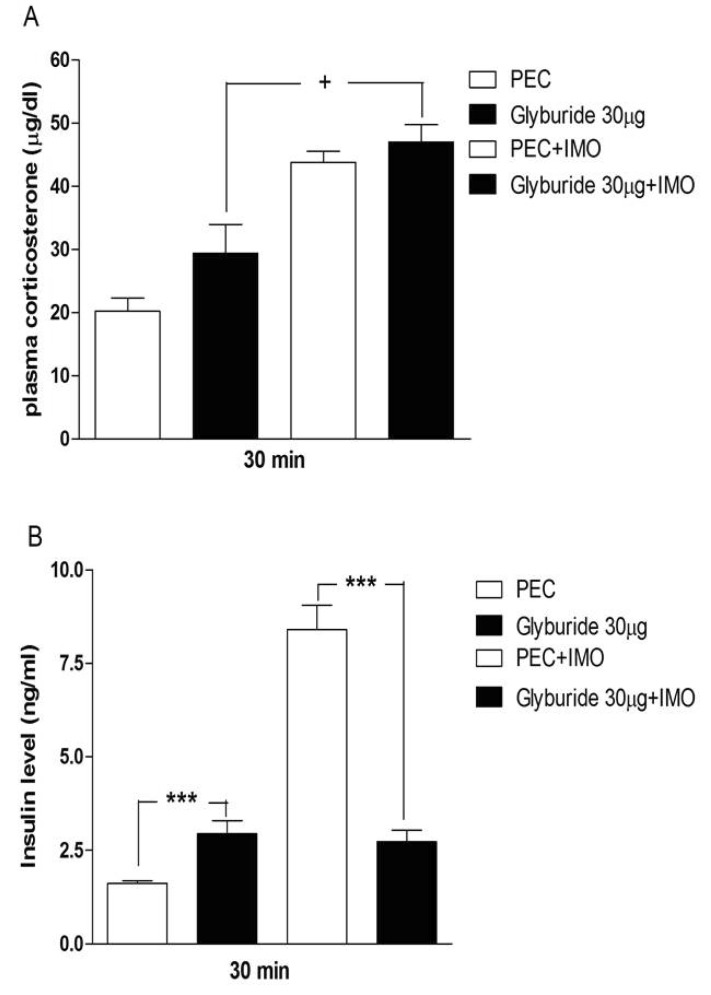
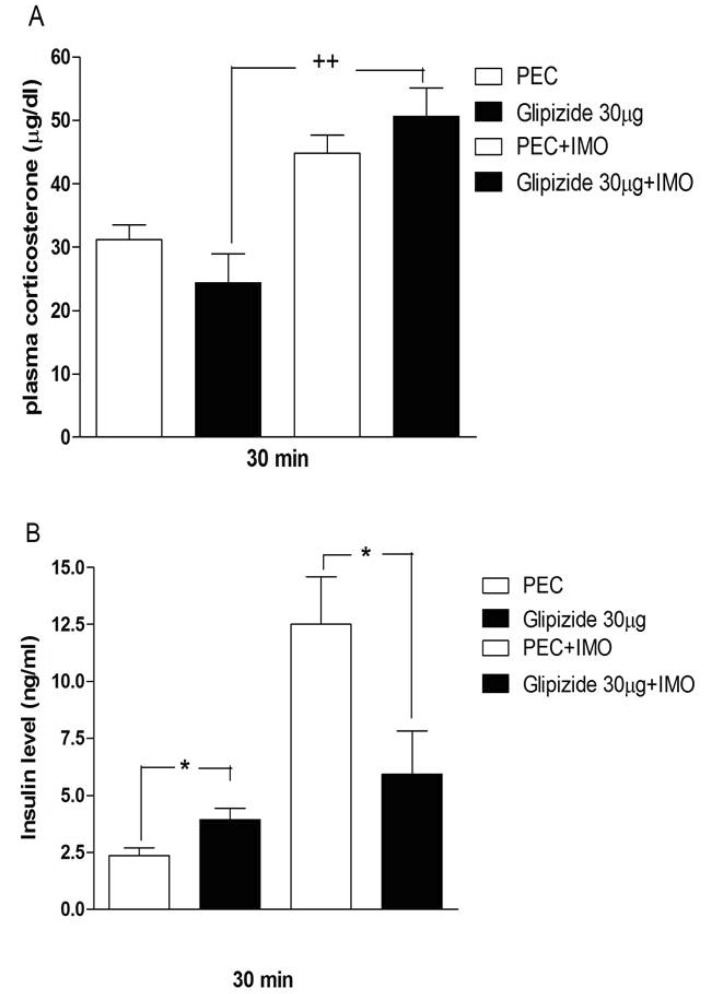
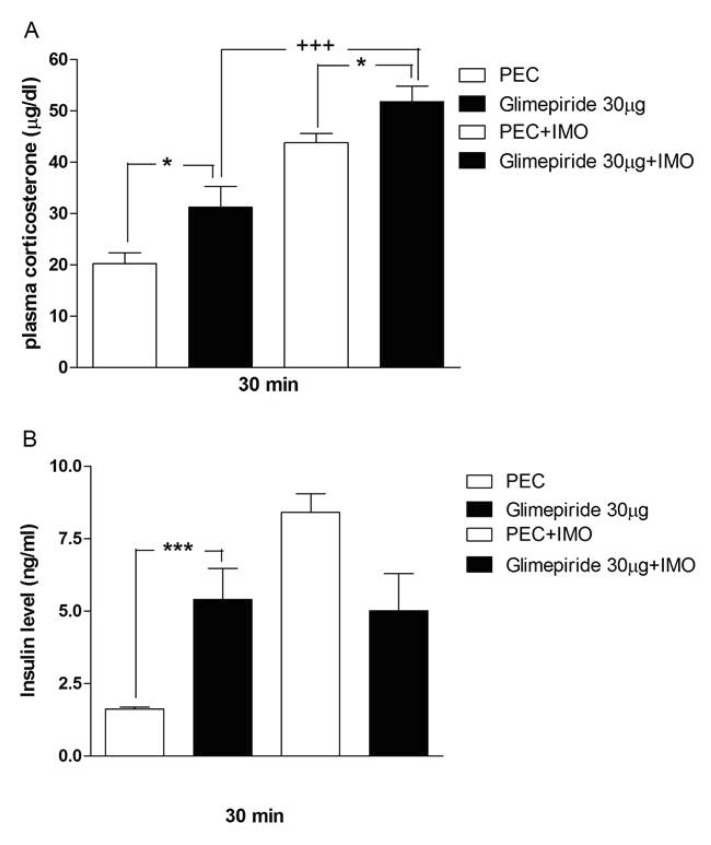
Effects of sulfonylureas pretreated i.c.v. on the blood corticosterone and insulin levels in immobilization stress model
Mice were pretreated i.c.v. with 30 µg of tolazamide, glyburide, glipizide or glimepiride for 10 min. Then mice were enforced into immobilization stress for 30 min and returned to the cage. The blood corticosterone and insulin levels were measured at 30 min after immobilization stress initiated. As shown in Figs. 2A, 3A, 4A and 5A, immobilization stress caused an elevation of plasma corticosterone level. In addition, tolazamide, glyburide, glipizide and glimepiride pretreated i.c.v. caused a further enhancement of the blood corticosterone level induced by immobilization stress. As shown in Figs. 2B, 3B, 4B and 5B, immobilization stress caused an elevation of plasma insulin level. In addition, tolazamide, glyburide, glipizide and glimepiride also increased plasma insulin level. However, tolazamide, glyburide, glipizide and glimepiride pretreated i.c.v. caused attenuated the blood insulin level induced by immobilization stress.
DISCUSSION
In the present study, the possible inhibitory action of sulfonylureas administered supraspinally against the elevation of the blood glucose level induced by immobilization stress was studied in ICR mice. We found in the present study that all of the second-generation sulfonylureas administered supraspinally exerts an inhibitory action against the increased blood glucose level induced by immobilization stress. However, first-generation sulfonylureas, except tolazamide, administered supraspinally does not alter the blood glucose level in immobilization stress model. These results suggest that some sulfonylureas exert their hypoglycemic effects at least by acting directly on the brain regions. Furthermore, from the findings of the present study, it can be speculated that, in addition to the direct action on pancreatic βcells, orally administered tolazamide, glyburide, glipizide and glimepiride may exert their anti-diabetic effects, at least, via acting on the brain. Although the exact reasons for differential actions of first-generation and second-generation sulfonylureas against immobilization stress-induced hyperglycemic effect are not currently known, earlier several studies have suggested that a second generation of sulfonylureas is far more potent than the first generation of sulfonylureas [12].
Several studies have demonstrated that the stress may cause increase in blood corticosterone level [21,22,23], this may cause increase in blood glucose level to meet energy demand during stress. Magariños et al. [24] demonstrated that, the stress-induced rise in plasma glucose may be attributed to corticosterone mediated enhanced metabolism to meet the increased demands of the body organs during stress. This elevation in corticosterone in stress condition may cause stress-induced hyperglycemia. In the present study, we found that the corticosterone level was significantly enhanced after 30 minutes of immobilization stress. In addition to this immobilization stress-induced corticosterone level is further increased by the i.c.v administration of sulfonylureas. These findings indicate that the corticosterone level increased by immobilization stress is just to supply the increased demand of energy to the organs during stress. Our results suggest that corticosterone appears not to be involved in the attenuation of blood glucose level by sulfonylureas. However corticosterone may help in blood glucose homeostasis against lowering effect of sulfonylureas.
Further, we examined whether insulin mediated pathway is involved in regulation of blood glucose level by i.c.v treatment with sulfonylureas. Ricart-Jané et al. [25] found that, a single immobilization stress application significantly increases the circulating level of corticosterone, glucose, insulin, glycerol, and ketone bodies, which is the typical response to acute stress. We found in the present study that the increase in blood glucose after acute immobilization stress is accompanied by an increase in plasma insulin level. The elevation of insulin level appears to maintain a homeostatic regulation of the blood glucose in immobilization stress. Sulfonylureas exert their anti-diabetic pharmacological effects in type 2 diabetes mellitus by improving glucose tolerance, stimulating insulin secretion peripheral tissues [26]. At in vitro, it is well known that sulfonylureas appear to block ATP-sensitive K+ channels at nanomolar concentrations in β-cells [27]. Sulfonylurea sensitive K+ channels inhibition lead to inhibition of gamma-aminobutyric acid (GABA) release, resulting in increase of insulin secretion [28,29]. We found in the present study that sulfonylureas administered supraspinally causes an elevation of plasma insulin level, suggesting that the activation of supraspinally located ATP-sensitive K+ channels may be responsible for an elevation of plasma insulin level. Furthermore, in a current study, we found that sulfonylureas administered supraspinally increase the blood insulin level. However, immobilization stress-induced insulin is attenuated by the supraspinal administration of sulfonylureas. Thus, it is speculated that the attenuation of the insulin level by sulfonylureas in stress model may be primarily due to the reduction of immobilization stress-induced blood glucose level by sulfonylureas.
We conclude in the present study that the supraspinal treatment with sulfonylureas attenuates the immobilization stress induced blood glucose level. Moreover, we found that corticosterone and insulin levels were enhanced by immobilization stress. In addition, immobilization stress-induced insulin level was attenuated by the supraspinal pretreatment with sulfonylureas, whereas immobilization stress-induced corticosterone level is further enhanced by sulfonylureas, suggesting that lowering effect of sulfonylureas administered supraspinally against immobilization stress-induced increase of the blood glucose level appears to be primarily mediated via elevation of the plasma insulin level.
 XML Download
XML Download