

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
KRGE
Korean Red Ginseng Extract
ROS
reactive oxygen species
eNOS
endothelial nitric oxide synthase
NADPH
nicotinamide adenine dinucleotide phosphate
DAF-FM
4-Amino-5-methylamino)-2', 7'-difluorofluorescein diacetate
DHE
dihydroethidine
PE
phenylephrine
ODQ
1H-[1,2,4]Oxadiazolo [4,3-a] quinoxalin-1-one
Ach
acetylcholine
TBA
Thiobarbituric acid
SNP
sodium nitroprusside
INTRODUCTION
Cardiovascular disease is the primary cause of morbidity and mortality in both industrialized and developing countries [1]. The occurrence of cardiovascular disease is going to be increased as the people ages, although successful treatments for cardiovascular risk factors, such as hypertension, hypercholesterolemia, and atherosclerosis, are rapidly developing.
The hallmark of the aging-related cardiovascular system is a decrease in nitric oxide (NO) bioavailability [2,3] and an increase in reactive oxygen species (ROS) production [4,5]. Superoxide (O2
·-) is a free radical that decreases NO bioavailability by rapidly scavenges NO. NO and O2
·- also can react to generate peroxynitrite (ONOO-), a highly oxidant molecule to macromolecules such as protein and DNA [6]. Simultaneous generation of NO and O2
·- can raise ONOO- to levels potentially harmful to homeostasis of vascular cell [6], and the imbalance of nitroso-redox system contributes to age-related endothelial dysfunction and vascular stiffness [7].
Nitric oxide (NO) is a major factor in endothelium-dependent vasorelaxation of the vascular system. NO act as a potent vasoprotective molecule by regulating vasoreactivity, inhibiting platelet activation and leukocyte adhesion, and preventing smooth muscle cell proliferation and migration in vasculature. NO is generated from its substrate L-arginine by eNOS enzyme in the endothelium and plays critical roles in the maintenance of vascular homeostasis. Arginase shares a common substrate L-arginine with eNOS and hydrolyzes L-arginine to L-ornithine and urea at the last step of the urea cycle. It is well documented that arginase reciprocally regulates NOS activity by modulating intracellular L-arginine bioavailability [7]. Thus, the balance of activities between these enzymes, arginase and eNOS, reciprocally regulates endothelial NO production.
Korean Red Ginseng water extract (KRGE) has been extensively studied, and its consumption has progressively increased in the general population. Ginseng has beneficial effects for treating various diseases including thrombosis, hyperlipidemia, cancer, and atherosclerosis [8,9,10,11,12]. KRGE exerts vasoprotective effects by eliminating O2
·- derived from NADPH oxidase [13], promoting endothelial cell proliferation and protecting against H2O2-dependent cell death [9,14], and inducing heme oxygenase-1 expression [15]. Ginseng extract exerts a direct vasodilatory effect by releasing NO in an endothelium-dependent manner [16]. Ginseng may exert its effects by activating phosphoinositide 3-kinase (PI3K)/Akt signal transduction in endothelial cells, which stimulates eNOS to produce NO, thereby leading to vascular system improvements [17].
Age-dependent vascular changes have been investigated in humans and a number of other species [18]. However, the degree to which these dysregulated mechanisms contribute to age-related vascular pathology remain to be elucidated. The contribution of vascular control mechanisms in healthy, aging, and disease conditions affected by vessel type and size [18]. In a previous study, we demonstrated that KRGE regulates NO production and enhances NO signaling in wild-type young mice [19]. The goal of this study was to investigate which mechanisms contribute to age-related endothelial dysfunction in aged mice and determine whether orally administered KRGE improves age-associated impairments in endothelial function.
METHODS
Materials
KRGE (Fig. 1, solid extract 64% from 6-year old ginseng roots, Rg1, 1.7 mg/g, Rb1 8.4 mg/g, number of product, 2020013) was obtained from Korea Ginseng Corporation (Daejeon, Korea) and directly dissolved in distilled water. Mn(III) Tetra (4-benzoic acid) porphyrin chloride (MnTBAP) and NG-nitro-L-arginine methyl ester (L-NAME) were purchased from Calbiochem. All reagents were from Sigma unless otherwise stated.
Animal protocol
Twenty young (10±3 weeks, 17.7±1.5 g) and twenty aged (55±5 weeks, 29.3±1.2 g) male mice (C57BL/6J) were used for all experiments. Mice were housed at 23℃ under a 12-h light/12-h dark cycle. All animals had access to water and food (Nara Biotech.) ad libitum. KRGE was orally administered at a dose of 0, 10, or 20 mg/day for 4 weeks. Given that each mouse consumed approximately 10 ml water/day, this represented a daily dose of ~20 mg/mouse/day of KRGE. The study protocols were approved in accordance with the Guide for the Care and Use of Laboratory Animals (Institutional Review Board, Kangwon National University).
Arginase activity assay
Aorta tissue lysates were prepared in lysis buffer (50 mM Tris-HCl, pH 7.5, 0.1 mM EDTA, and protease inhibitors). Lysates were homogenized at 4℃ and centrifuged for 20 min at 14,000×g at 4℃. The supernatants were used to measurement of arginase activity as previously described [20].
Western blotting analysis
Aortic vessels from C57BL/6 mice were lyzed in buffer (50 mM Tris-HCl, 150 mM NaCl, 1% Nonidet P-40, 1 mM EDTA, protease inhibitors (Roche co.)) and centrifuged for 30 min at 14,000×g. The amount of protein in the supernatant was determined by the Bradford method. Proteins (100 µg) were loaded on 10% SDS-PAGE and then transferred to a nitrocellulose membrane (Bio-Rad). The blots were incubated with a polyclonal anti-arginase II (Santa Cruz), anti-endothelial nitric oxide synthase (eNOS, BD Bioscience), or anti-actin (Santa Cruz) antibodies followed by the corresponding secondary antibody (Amersham). The signals were detected using an enhanced chemiluminescence detection reagent with X-ray films.
Determination of eNOS dimerization
eNOS dimers and monomers were separated using low-temperature SDS-PAGE and visualized by western blot analysis with specific antiserum as previously described [21]. Band intensities were analyzed by NIH ImageJ Software.
Estimation of NO and ROS generation in isolated mice aorta
We used 4-amino-5-methylamino-2',7'-difluorescein diacetate (DAF-FM) to estimate NO production and dihydroethidine (DHE) to measure ROS production. The change in fluorescence intensity at different time intervals was measured using a microscope as described previously [22].
Aortic vascular tension assay
We measured vascular tension to determine whether increased NO production by KRGE contributes to vasorelaxation in aged mice. Mice were euthanized using isoflurane and the thoracic aorta was rapidly removed. The aorta was placed in ice-cold oxygenated Krebs-Ringer bicarbonate solution (118.3 mM NaCl, 4.7 mM KCl, 1.2 mM MgSO4, 1.2 mM KH2PO4, 1.6 mM CaCl2, 25 mM NaHCO3, 11.1 mM glucose) and cleared of adherent connective tissues. The mouse aorta was cut into 1.5-mm rings and suspended between two wire stirrups (150 µm) in a myograph (Multi myograph system DMT-620) immersed in 10 ml Krebsringer (95% O2 -5%CO2, pH 7.4, 37℃). One stirrup was connected to a three-dimensional micromanipulator and the other to a force transducer. The rings were passively stretched at 10-min intervals in 100-mg increments to reach optimal tone (600 mg). After the arterial rings were stretched to their optimal resting tone, the contractile response to 100 mM KCl was determined. For the constriction assay, phenylephrine (PE) was applied at different concentrations (10-9~10-5 M), and cumulative responses were recorded. For NO-dependent relaxation responses, vessels were preconstricted with PE (10-6 M) for 15 min, and the endothelium-dependent vasodilator acetylcholine (Ach, 10-9~10-5 M) was then added to determine relaxation activity. To further confirm the vasorelaxation activity we observed occurred in a NO-dependent manner, we added 1 µM 1H-[1,2,4]oxadizolo[4,3-a]quinoxalin-1-one (ODQ), an inhibitor of guanylate cyclase, at the end of the experiments.
Determination of thiobarbituric acid-reactive substances (TBARS)
The precipitate of plasma samples by mixing with trichloroacetic acid (20%) was dispersed in H2 SO4 (0.05 M). Thiobarbituric acid (TBA, 0.2% in 2 M sodium sulfate) was added and heated for 30 min in a boiling water bath. TBARS products extracted with n-butanol and absorbance at 532 nm was measured [23]. Malondialdehyde (MDA) was used as a standard.
Statistical Analysis
All data are represented as mean±S.D. of at least four independent experiments. An unpaired Student's t-test or 1-way ANOVA was used to assess significant differences. A value of p<0.05 was accepted as significant. Experimental data were analyzed with the GraphPad Prism program (GraphPad Software 4.02).
RESULTS
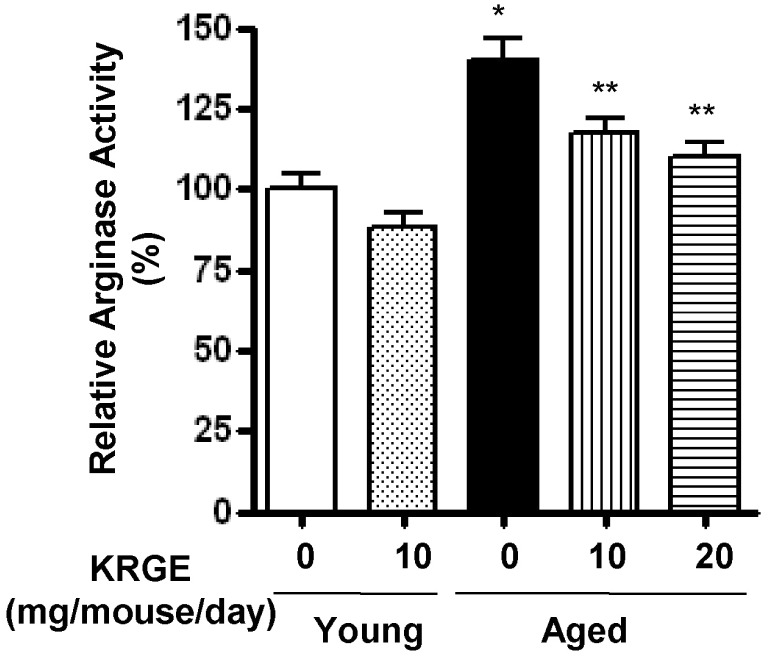
Effect of KRGE on arginase activity in aortas of aged mice
Mice were orally administered KRGE at 10 or 20 mg/mouse/day for 4 weeks. Arginase activity was significantly higher in the aorta of aged mice than in the young control mice (140.0±13.5 vs. 100±6.9%, p<0.01) (Fig. 2). KRGE doses reduced arginase activity in the aged mice to 117.1±10.2 (10 mg/mouse/day) and 110.0±9.2 (20 mg/mouse/day) (p<0.05).
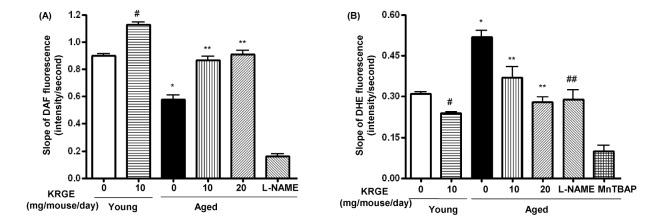
KRGE enhances NO generation and attenuates ROS production in endothelium of aged mice
Given that arginase competes with nitric oxide synthase (NOS) for L-arginine and negatively regulates NO production, we measured whether KRGE-dependent inhibition of arginase activity was associated with an increase in NO production. As demonstrated in Fig. 3A, NO production was significantly decreased in aged mice compared to young control mice (0.58±0.16 vs. 0.90±0.06 change in DAF fluorescence/sec, p<0.01). KRGE administration restored NO generation to 0.87±0.11 and 0.91±0.14 change in DAF fluorescence/sec in the 10 mg/mouse/day and 20 mg/mouse/day groups, respectively (p<0.01). KRGE also increased NO production in young mice (1.13±0.09 change in DAF fluorescence/sec, p<0.01). ROS generation was significantly higher in aged mice than in their young counterparts (Fig. 3B, 0.52±0.13 vs. 0.31±0.04 change in DHE fluorescence/sec, p<0.01), and was significantly reduced with 10 mg/mouse/day and 20 mg/mouse/day KRGE (0.37±0.13 and 0.28±0.06 change of DHE fluorescence/sec, respectively, p<0.01 compared to untreated aged mice). KRGE also attenuated ROS generation in young mice (0.24±0.02 change in DHE fluorescence/sec, p<0.01).Treatment with the NOS inhibitor L-NAME reduced ROS generation in aged mice (0.29±0.11 change in DHE fluorescence/sec, p<0.01).
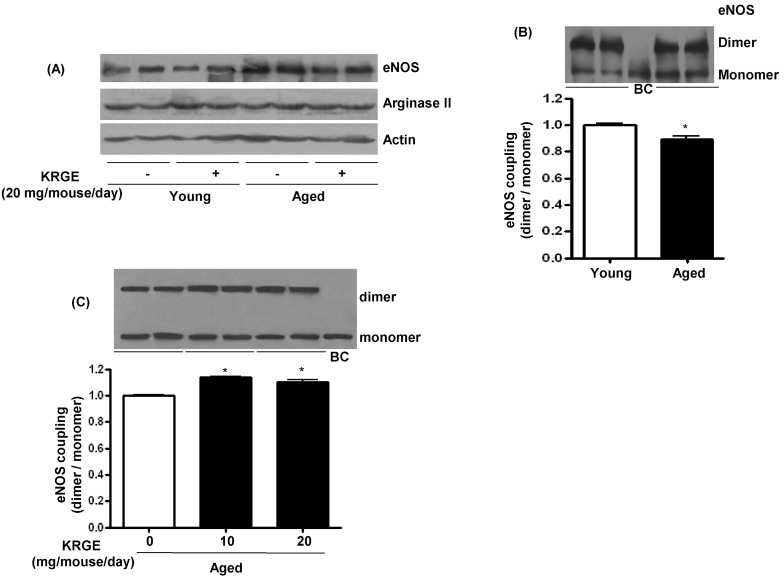
KRGE administration enhanced eNOS coupling
Based on the NO production and ROS generation data, we next assessed protein expression to find the underlying mechanism. The eNOS protein expression was significantly higher in aged mice than in young mice (1.23±0.04 vs. 1.0±0.03, p<0.01) and was unaffected by KRGE administration (Fig. 4A). Arginase II protein expression was not different between the groups (Fig. 4A). The eNOS coupling was lower in aged mice than in their young counterparts (Fig. 4B) and was restored with KRGE administration (Fig. 4C). These results imply that endothelial function is impaired through attenuated eNOS coupling despite increased eNOS expression, and that KRGE administration can increase eNOS coupling in the aorta of aged mice.
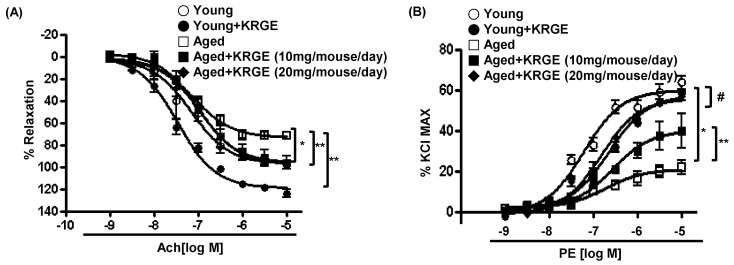
NO-dependent relaxation of aortic vessels was augmented with KRGE administration
With the above data that KGWE administration induced eNOS coupling and NO production, we measured vascular tension to determine whether increased NO production by KGWE contributes to vasorelaxation in aged mice. The vasorelaxation responses in aged mice were significantly attenuated compared to those from the young (Emax 73.30±2.50% vs. 97.16±4.08%, p<0.01; -logEC50 6.81±0.09 M vs. 7.23±0.13 M, *p<0.01). KRGE administration to young and aged mice enhanced vasorelaxation responses (**p<0.01) (Fig. 5A). As shown in Fig. 5B, the responses to PE were markedly attenuated in aortas from aged mice (Emax, aged vs. young, 20.93±1.68% vs. 59.92±1.83%, p<0.01; -log EC50, aged vs. young, 7.72±0.20 M vs. 8.21±0.08 M, *p<0.01). However, KRGE administration enhanced vasoconstrictor responses in aged mice as determined by Emax (40.06±3.32% and 55.84±1.54% for 10 mg/mouse/day and 20 mg/mouse/day KRGE, respectively, **p<0.01 vs. untreated aged), but not -logEC50 was not significant between groups. Interestingly, KRGE attenuated vasoconstriction response to PE in young mice (#p<0.05). The response to the endothelium-independent vasodilator sodium nitroprusside (SNP) was not significantly different in any of the groups (data not shown).
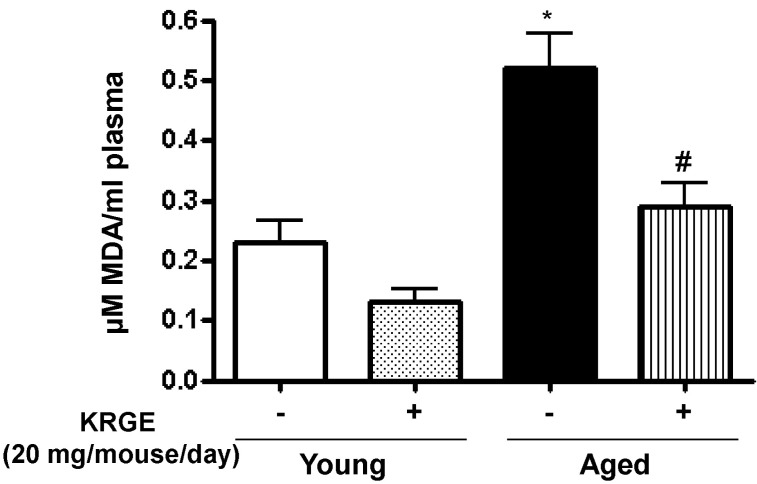
KRGE prevents peroxynitrite formation in aged mice
Because oxidative species such as peroxynitrite increase arginase activity [24], and enhanced peroxynitrite formation is associated with vascular aging [25], we measured lipid peroxidation using a TBARS assay. Lipid peroxidation was significantly increased in the plasma of aged mice (0.52±0.13 vs. 0.23±0.08 µM MDA/ml plasma in young mice, p<0.01) (Fig. 6). KRGE administration (20 mg/mouse/day) reduced lipid peroxidation in aged mice to a level similar to young untreated mice (0.29±0.09 µM MDA/ml plasma, p<0.01 compared to aged untreated mice) (Fig. 6).
DISCUSSION
Endothelial arginase reduces eNOS activity by depleting the critical substrate L-arginine, and this increased arginase activity reduces NO bioavailability and contributes to vascular diseases such as aging, hypertension, and atherosclerosis [7,21,26,27]. Here we showed that oral administration of KRGE for 4 weeks increased NO generation, reduced ROS production by inhibiting arginase activity, induced vasorelaxation in aortic vessels from aged mice, and reduced plasma peroxynitrite formation. These findings suggest KRGE consumption may provide vasoprotective benefits for the aging cardiovascular system.
Ginseng is beneficial in the treatment of various diseases. KRGE eliminates O2
·- [13], promotes endothelial cell proliferation [9], protects against H2O2-induced cell death by [14], and induces heme oxygenase-1 expression [15] in the vasculature. We demonstrated that KG-WE-dependent arginase inhibition contributed to an increase of NO generation and decease of ROS production in isolated mice aortas (Fig. 3). These effects resulted in the induction of NO-dependent vasorelaxation (Fig. 5). These results are consistent with previous demonstrations that arginase inhibition accentuates NO release in rat aortic endothelium [7], bovine pulmonary endothelial cells [28], and porcine coronary artery model [29]. The main mechanism of KG-WE-dependent NO release is likely due to increased eNOS coupling resulting from increased L-arginine availability (Fig. 4), in line with previous data [21]. Therefore, these data further support that KRGE can play important role in improving age-related vascular health. The beneficial effects of ginseng on the vascular system are likely complex and may depend on the activation of Akt/PI3K signal transduction [17], inhibition of angiotensin converting enzyme [30], and inhibition of calcium ion influx [31].
Aging is associated with changes in arterial wall structure and function, the most frequent being luminal enlargement, vessel wall thickening due to intimal and medial expansion, elastin depletion and fragmentation, collagen and calcium deposition, glycation of proteins, and impaired vasomotor function associated with endothelial dysfunction [32,33,34]. These alterations increase vascular stiffness, which is an independent risk factor for cardiovascular morbidity and mortality [35,36,37]. Arginase increases the synthesis of polyamines, which stimulate vascular smooth muscle and endothelial cell proliferation, and L-proline, which increases collagen deposition. Together, these cellular alterations contribute to endothelial dysfunction and arterial stiffening associated with aging. Although increased arginases activities were well associated with endothelial dysfunction in diseases-related animal models [7,28,29], arginase itself currently had a limitation of usefulness to extent the human in terms of aging.
Vascular ROS production is enhanced in aged blood vessels [4,25,38,39], and the enzymatic system eNOS may contribute to increased ROS production in pathophysiological states. Typically, eNOS produces the vasoprotective molecule NO. However, it can also produce O2
·- in the absence of substrate, L-arginine, or cofactor, BH4, because electrons flow from the reductase domain in the heme to molecular oxygen rather than to L-arginine in uncoupled eNOS. We showed increased eNOS uncoupling (Fig. 4B), arginase activity (Fig. 2), and peroxynitrite formation (Fig. 6) in aged vessels. Furthermore, preincubation of L-NAME, the eNOS inhibitor, attenuated ROS production on the endothelium of aged mice (Fig. 3B), which is consistent with previous observations [40]. Peroxynitrite may stimulate arginase II activation via ROCK-dependent pathway without a change in mRNA and protein levels [24]. Peroxynitrite reacts also with the redox-active cysteine (Cys18) of RhoA, which results in enhanced release of GDP from RhoA and thus modulates its activity [4]. Indeed, we found plasma peroxynitrite formation, as measured by the TBARS assay, was significantly higher in aged mice compared to their young counterparts (Fig. 6), which is consistent with previous research [25].
The promoter region of the eNOS gene has binding site to a shear stress-responsive element [41] indicates that shear stress and hemodynamic forces may increase eNOS expression [42,43]. Consistent with these findings, eNOS expression is higher in the aorta of aged animals [44], but is not changed in the artery [45] and is decreased in the arteriole [3]. Increased eNOS protein expression in the aged aorta may be a mechanism to compensatory the endothelial dysfunction associated with increased arginase activity (Fig. 4A).
The expression of inducible nitric oxide synthase (iNOS) has been reported in old rat model [46]. Therefore, increased iNOS expression and/or activity may be an additional possible mechanism to explain the decreased contraction in the aortic rings of aged mice in response to PE (Fig. 5B). Although we could not detect the expression of iNOS protein at arotic vessels from aged mice in the presence or absence of KRGE (data not shown), we could not exclude the involvement of iNOS.
In summary, we demonstrate that KGRE inhibits the age-associated increase in arginase activity in the aorta, and is associated with enhanced eNOS dimerization and increased NO production. Furthermore, KRGE augments vasorelaxation in a NO-dependent manner and attenuates peroxynitrite formation. These findings suggest that KGRE possesses therapeutic potential for cardiovascular diseases associated with aging.
 XML Download
XML Download