

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Asthma is a chronic disease characterized by airway inflammation, mucus hyperproduction and bronchial hyperreactivity.1 Majority of asthma is triggered by immune responses to inhaled allergens such as house dust mite (HDM), microbes, pollutants and tobacco smoke. Asthma is a complex and heterogeneous disease with different etiologies. Although it is well established that dysregulated T helper (Th) type 2 immunity is the main cause of asthma,234 some patients have Th17-mediated airway inflammation with increased airway neutrophils.5 Studies show that interleukin (IL)-17A is markedly elevated in patients with moderate to severe asthma and positively correlated with the disease severity.67 Similarly, IL-17 production and neutrophil recruitment are increased in the airways of mice after HDM sensitization and challenge.8
The transcription factor B-cell chronic lymphocytic leukemia/lymphoma 11B gene (Bcl11b) is broadly expressed in all T cell subsets and plays a critical role in T-cell development, differentiation, function and survival. The deletion of Bcl11b in progenitors blocks T-cell commitment at the double negative 2 (DN2) stage and impairs positive selection of CD4 and CD8 single positive (SP) thymocytes and survival of double-positive (DP) thymocytes.91011 Bcl11b also appears to function predominantly as a transcription factor in mature T cells including Th cells, cytotoxic T cells (CTLs), regulatory T (Treg) cells, γδ T cells and iNKT cells.1213 The role of Bcl11b in T-cell development has been defined, but the function of Bcl11b in the immune response is still poorly understood. Bcl11b suppresses skin inflammatory responses by regulating the expression of Th2-type cytokines14 and restricts plasticity of Th17 cells by blocking the expression of Th2 program.15 We previously demonstrated that Bcl11b can increase the proliferative ability of human Th subsets.16 However, how Bcl11b regulates different Th subsets is unknown and whether Bcl11b is important in Th cell differentiation remains to be determined.
Our previous experiments suggested that alteration in Bcl11b expression led to a clear dysregulation of the transforming growth factor (TGF)/Smad signaling pathway in naïve T cells. Since the TGF/Smad signaling pathway is essential for Th17-cell differentiation, in this study, we analyze the expression of Bcl11b in both asthmatic patients and HDM-induced mouse model of asthma. We describe for the first time that Bcl11b expression is negatively correlated with IL-17A and TGF-β1 in patients with severe asthma (SA). We also find that Bcl11b negatively regulates the TGF-β1/Smad pathway to control IL-17A production.
MATERIALS AND METHODS
Human subjects and samples preparation
Twenty-five patients with mild asthma (MA) and 15 patients with SA at the Department of Pulmonary Medicine, The First Affiliated Hospital of Shenzhen University (Shenzhen, China) were recruited into this study. The diagnosis of asthma and the assessment of its severity were carried out according to the Global Initiative for Asthma (GINA) and ERS/ATS guidelines. All participants were subjected to forced expiratory volume in the first second (FEV1) test and total IgE and HDM-specific IgE measurements. Patients with MA were collected at the time of diagnosis in outpatient, before administration of asthma medication. Patients with SA were hospitalized patients who showed exacerbation in symptoms such as wheezing, breathlessness and chest tightness 24 hours prior to admission to the emergency department and received only rescue medication. Blood samples from these patients were obtained after admission to the emergency department but before the initiation of inhaled corticosteroids (ICSs)/long-acting β2-agonists (LABAs) treatment. Then, these patients were treated with therapy (medications for GINA steps 4–5 asthma). Fifteen healthy donors, with normal pulmonary function and negative allergy tests, were selected as normal controls (NCs). The study was approved by the Research Ethics Board of School of Medicine, Shenzhen University (Shenzhen, China). Written informed consent was obtained from all individuals enrolled in the study.
Heparinized peripheral venous blood (4–5 mL) was collected from each participant. Peripheral blood mononuclear cells (PBMCs) were separated immediately using the Ficoll-Hypaque gradient centrifugation method. CD4+ T cells were isolated from PBMCs by human CD4+ Micro Beads (Miltenyi Biotec, Bergisch Gladbach, Germany).
Animals
Female BALB/c mice (6–8 weeks) were purchased from the Animal Center of Guangdong Province. The mice were maintained in a specific pathogen-free facility at the Experimental Animal Center of Shenzhen University. All animal care and experimental protocols were carried out in accordance with the Institutional Guidelines for Care and Use of Laboratory Animals at Shenzhen University.
HDM-induced allergic airway inflammation
The antigen sensitization and challenge in the murine model of allergic asthma were performed as previously described,1718 and the scheme of the study is shown in Fig. 1A. Briefly, BALB/c mice received immunization with 100 µg of HDM extract in 100 µL of PBS with 2% aluminum hydroxide by intraperitoneal injection on days 0, 3, and 7. From days 15 to 21, the mice were challenged intranasally with 50 µg of HDM in 50 µL of PBS. The PBS control group was treated with PBS in all procedures. Twenty-four hours after the final challenge, airway hyperreactivity (AHR) was assayed and on day 23 all the mice were sacrificed. Purified mouse CD4+ T cells of both the HDM and PBS groups were isolated from spleens by mouse CD4+ Micro Beads (Miltenyi Biotec).
Fig. 1
Levels of Th2 and Th17 cytokines in HDM-induced mouse model of asthma. (A) The design of the HDM-induced mouse model of asthma. (B) The numbers of lymphocytes, eosinophils, neutrophils and macrophages in the BALF were evaluated 48 hours after the last HDM challenge. (C) The ARH to Mch were measured 24 hours after the last HDM challenge. (D) The levels of HDM-specific IgE and IgG1 in BALF were measured by ELISA. (E) IL-4, IL-5, and IL-13 levels in BALF were quantified by ELISA. (F) IL-17A and TGF-β1 levels in BALF were measured by ELISA. The results are representative of 3 independent experiments (n = 3–5).
Data represent mean ± standard error of the mean.
Th, T helper; HDM, house dust mite; BALF, bronchoalveolar lavage fluid; AHR, airway hyperreactivity; Mch, methacholine; Ig, immunoglobulin; ELISA, enzyme-linked immunosorbent assay; IL, interleukin; ns, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.

AHR measurement
AHR to methacholine (Mch) aerosol was evaluated using unrestrained whole-body plethysmography with a 4-chamber system (Buxco Research Systems, Wilmington, NC, USA). Firstly, the baseline response was recorded for 5 minutes at steady respiration after 10-minute acclimation. Then, the mice were subjected to Mch inhalation at increasing concentrations (6.25, 12.5, 25, and 50 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) or PBS. Tests were temporally separated to allow the respiratory intensity to drop back to baseline. The percentage curves for Penh values at different Mch doses were plotted.
Bronchoalveolar lavage (BAL)
BAL was performed as previously described.18 The supernatant was collected for cytokine determination by enzyme-linked immunosorbent assay (ELISA). Pellets were resuspended and counted with hemocytometer. Cytospin preparations were stained manually with Wright-Giemsa stain. The percentages of lymphocytes, eosinophils, neutrophils and macrophages were determined by counting at least 200 leukocytes in randomly selected areas under a light microscope.
Cell culture and transfection
Splenocytes isolated from spleens of BALB/c mice were first challenged with HDM (30 µg/mL) overnight. Then, the cells were divided into 4 groups (1 × 107 cells/group) and treated as follows: 1) pI2 (pIRES2-EGFP vector control), 2) pBcl11b (pIRES2-EGFP-Bcl11b recombinant plasmid group), 3) TGF-β1 (TGF-β1 stimulation group), and 4) pBcl11b+TGF-β1 (pBcl11b + TGF-β1 treatment group). To up-regulate Bcl11b expression in vitro, cells were transfected with 4 µg of mouse Bcl11b plasmid (GeneChem, Shanghai, China) using Amaxa Nucleofector Kit (Lonza, Basel, Switzerland). For TGF-β1 treatment, recombinant TGF-β1 (Peprotech, Rocky Hill, NJ, USA) was prepared as a stock solution at a concentration of 0.1 mg/mL in 10 mM citric acid. Cells were cultured with 5 µM TGF-β1 in the presence (pBcl11b + TGF-β1 group) or absence (TGF-β1 group) of pBcl11b. After 3 days of culture, the levels of cytokines in the supernatant were measured by ELISA and the cells were collected to detect gene expression by quantitative real-time (qRT)-polymerase chain reaction (PCR) and Western blotting.
qRT-PCR
Total RNA was isolated using Trizol Reagent (Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized using the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA). Quantification of human Bcl11b mRNA was performed as previously described.16 Bcl11b copies were normalized to 105 copies of the β2m reference gene. Changes in the mRNA relative expression of human and mouse IL-17A and TGF-β1, and mouse Bcl11b, Smad2, Smad3, and Smad4 were determined by SYBR Green based real-time PCR. Prime and probe sequences are detailed in Table 1. PCR amplification was performed in a total volume of 20 µL with 1 µL of cDNA, 0.5 mmol/L of each primer and 9 µL of SYBR Green I Master Mix (Takara, Beijing, China). Relative target gene expression was calculated by normalization to β-2M or β-actin with the 2−ΔΔCt method and presented as folds of change against controls.
Table 1
Prime and probe sequences

ELISA
The levels of human and mouse cytokines were determined by ELISA according to the instructions of reagent kits.
Western blotting
Proteins were extracted from cells, separated by SDS-PAGE and transferred to PVDF membranes. After blocking, the membranes were incubated overnight with antibodies against Bcl11b, Smad2, phosphorylated (p)-Smad2, Smad3, p-Smad3, Smad4, and β-actin (Cell Signaling Technology, Danvers, MA, USA). Immune reaction bands were detected by using specific secondary antibodies (Cell Signaling Technology) and enhanced chemiluminescence reagents (EMD Millipore, Billerica, MA, USA), and viewed by Kodak Image Station 4000MM (Eastman Kodak, Rochester, NY, USA). The relative abundance of protein was determined by quantitative densitometry using Image J software (National Institutes of Health, Bethesda, MA, USA), and the data were normalized to β-actin.
Statistical analysis
Data are expressed as mean ± standard error of the mean of at least 3 independent experiments. Statistical analysis was performed using Prism 7.0 (Graphpad Software, La Jolla, CA, USA). A 2-tailed Student's t-test was used for pairwise comparison. Where appropriate, the ANOVA test and the Bonferroni post hoc test were used to analyze multiple comparisons and the Mann-Whitney U test was used to analyze non-parametric data. The correlations were examined by Spearman's rank correlation test. The P values of < 0.05 were considered significant.
RESULTS
Bcl11b expression is negatively correlated with the level of IL-17A or TGF-β1 in SA patients
According to the GINA guideline, patients may be divided into 2 groups: MA and SA. No significant differences were identified in terms of age, sex and BMI between the MA, SA and NC groups. We observed significant increases in blood absolute eosinophil/neutrophil count, total IgE and HDM-specific IgE in SA group in comparison to the MA group. FEV1 (% pred.) was lower in patients with MA or SA than that in NC (Table 2).
Table 2
Demographic and characteristics of human subjects

To determine whether Th2 cytokines (IL-4, IL-5, and IL-13) and Th17 cytokine (IL-17A) are both implicated in asthmatic patients positive for HDM-specific IgE, we measured the plasma levels of IL-4, IL-5, IL-13, and IL-17A. We found that IL-4, IL-5, and IL-13 expression were increased in the MA and SA groups when compared to NC. The levels of IL-4 and IL-5 were higher in the SA group than in the MA group; however, no significant difference was observed in the levels of IL-13 between these 2 groups (Fig. 2A). Patients in the SA group showed an increased concentration of IL-17A compared with the other groups. The levels of IL-17A in MA and NC were no different (Fig. 2B). In addition, we noted that the levels of TGF-β1 were increased in both the MA and SA groups compared to NC (Fig. 2B).
Fig. 2
Expression levels of Bcl11b, Th2 and Th17 cytokines from patients with asthma. (A) IL-4, IL-5, and IL-13 levels in the plasma of patients with MA (n = 25), SA (n = 15) and NC (n = 15) were quantified by ELISA. (B) IL-17A and TGF-β1 levels in the plasma of MA, SA and NC were measured by ELISA. (C) The mRNA copies of Bcl11b in purified CD4+ T cells were normalized by 105 β2M using TaqMan based real-time PCR. (D) The relative mRNA expression of IL-17A and TGF-β1 in purified CD4+ T were normalized by the 2−ΔΔCt method using SYBR GREEN based real-time PCR. (E) The correlation between Bcl11b copies and relative IL-17A mRNA levels from patients with SA. (F) The correlation between Bcl11b copies and relative TGF-β1 mRNA levels from patients with SA.
Data represent mean ± standard error of the mean.
Th, T helper; IL, interleukin; MA, mild asthma; SA, severe asthma; NC, normal controls; ELISA, enzyme-linked immunosorbent assay; TGD, transforming growth factor; PCR, polymerase chain reaction; ns, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.

Our previous study demonstrated a negative impact of Bcl11b on TGF-β/Smad signaling in naïve T cells using global gene expression analysis. We hypothesize that a reduction of Bcl11b results in enhanced TGF-β/Smad signaling that is essential for Th17 differentiation and IL-17A expression. We examined Bcl11b mRNA levels by TaqMan-based real-time PCR in CD4+ T cells, and found that Bcl11b copies were significantly decreased in the MA or SA group compared with NC donors (Fig. 2C). Conversely, IL-17A and TGF-β1 relative mRNA levels were increased (Fig. 2D). However, there were no differences in Bcl11b, IL-17A, and TGF-β1 mRNA expression between the MA and SA groups. We also analyzed the possible correlation between Bcl11b and Th2 cytokines/IL-17A/TGF-β1. In the SA group, the copies of Bcl11b were inversely correlated with IL-17A and TGF-β1 mRNA levels (Fig. 2E and F). However, in the MA group, no significant correlations were found between IL-17A or TGF-β levels and the copies of Bcl11b (data not shown). In addition, there were no differences in the correlations of Th2 inflammation markers (IL-4, IL-5, and IL-13) and Bcl11b between the MA and SA groups (data not shown). Together, these findings suggest that Bcl11b regulates IL-17 production in asthmatic patients positive for HDM-specific IgE.
Bcl11b expression is down-regulated, but TGFβ/Smad signaling and IL-17 expression are enhanced in the HDM-induced mouse model of asthma
To explore the role of Bcl11b in the pathogenesis of asthma, we established a HDM-induced allergic airway inflammation model (Fig. 1A). We found that mice immunized and challenged with HDM exhibited increased inflammatory cells (lymphocytes, eosinophils, neutrophils, and macrophages) in bronchoalveolar lavage (Fig. 1B) and AHR (Fig. 1C). Consistently, HDM-specific IgE and IgG1 (Fig. 1D), Th2 cytokines (IL-4, IL-5, and IL-13) (Fig. 1E), IL-17A and TGF-β1 in BALF (Fig. 1F) were all significantly increased in mice immunized and challenged with HDM. These results indicated that HDM-challenged mice appear to have both Th2- and Th17-mediated allergic inflammation.
To determine the mechanism of Bcl11b regulating IL-17 in asthma, we assessed expression of Bcl11b, IL-17A, TGF-β1, Smad2, Smad3, and Smad4 in CD4+ T cells using qRT-PCR or western blotting. CD4+ T cells isolated from HDM-treated mice showed a 2.56-fold decrease in Bcl11b mRNA, but significantly up-regulated IL-17A and TGF-β1 mRNA compared with control mice (Fig. 3A). Western blot analysis showed similar change in Bcl11b protein expression (Fig. 3B and C). Although the total protein amounts of Smad2, Smad3, and Smad4 did not significantly differ between the 2 groups, the levels of phosphorylated Smad2 and Smad3 were significantly increased in CD4+ T cells from HDM-treated mice (Fig. 3B and C). Taken together, our data suggest that down-regulation of Bcl11b in response to HDM treatment may enhance TGF-β/Smad signaling in CD4+ T cells to promote IL-17A expression.
Fig. 3
Expression of Bcl11b, IL-17A and the TGF-β/Smad signaling pathway in HDM-treated mice. (A) The relative mRNA expression of Bcl11b, IL-17A and TGF-β1 in CD4+ T cells were normalized by 2−ΔΔCt method using SYBR GREEN based real-time PCR. (B) The protein expression of Bcl11b, Smad2, pSmad2, Smad3, pSmad3, and Smad4 in CD4+ T were determined by western Blot. (C) Quantification of western blotting results. Band intensities were measured using Image J. The results are representative of 3 independent experiments (n = 3–5).
Data represent mean ± standard error of the mean.
IL, interleukin; TGF, transforming growth factor; HDM, house dust mite; PCR, polymerase chain reaction; ns, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.

Bcl11b negatively regulates IL-17A production through the TGFβ/Smad2 pathway
Our data demonstrated an inverse relationship between Bcl11b expression and Smad2/3 phosphorylation or IL-17A accumulation. To confirm whether Bcl11b negatively regulates the TGF-β/Smad pathway, we transfected HDM-stimulated mouse splenocytes with mouse Bcl11b expression plasmid in the presence or absence of TGF-β1. Compared with the pI2-empty vector group, the pBcl11b-transfected groups showed increased Bcl11b expression at both mRNA (Fig. 4A) and protein levels (Fig. 4E and F). Ectopic expression of Bcl11b suppressed mRNA expression of IL-17A, TGF-β1, and Smad2, but did not affect Smad3 expression (Fig. 4B and C). Down-regulation of IL-17A by Bcl11b overexpression was confirmed in the culture supernatant by ELISA (Fig. 4D). In addition, Western blot analysis demonstrated that overexpression of Bcl11b decreased Smad2 activation, evidenced by a 1.3-fold reduction in Smad2 phosphorylation (Fig. 4E and F).
Fig. 4
Effect of Bcl11b on TGF-β1/Smad signaling and IL-17 expression in vitro. (A) The relative mRNA expression of Bcl11b in HDM-cultured splenocytes from naïve BALB/c mice were normalized by 2−ΔΔCt method using SYBR GREEN based real-time PCR (pI2: pIRES2-EGFP vetor control; pBcl11b: pIRES2-EGFP-Bcl11b plasmid; TGF-β1: TGF-β1 stimulation; pBcl11b + TGF-β1: pBcl11b + TGF-β1 treatment). (B) The relative mRNA expression of IL-17A and TGF-β1 in cultured lymphocytes were normalized by 2−ΔΔCt method using SYBR GREEN based real-time PCR. (C) The relative mRNA expression of Smad2 and Smad3 in cultured cells were normalized by 2−ΔΔCt method using SYBR GREEN based real-time PCR. (D) IL-17A levels in the supernatant were measured by ELISA. (E) The protein expression of Bcl11b, Smad2, pSmad2, Smad3, and pSmad3 in cultured cells were determined by western Blot. (F) Quantification of western Blot results. Band intensities were measured using Image J. The results are representative of the 3 independent experiments (n = 3–5).
Data represent mean ± standard error of the mean.
TGF, transforming growth factor; HDM, house dust mite; PCR, polymerase chain reaction; ELISA, enzyme-linked immunosorbent assay; ns, not significant.
*P < 0.05, †P < 0.01, ‡P < 0.001.
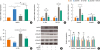
We finally investigated whether Bcl11b also suppresses TGF-β1-stimulated Smad activation and IL-17A expression. While TGF-β1 treatment enhanced IL-17A expression at both mRNA and protein levels, such effects were abrogated by ectopic expression of Bcl11b (Fig. 4B and D). Accordingly, TGF-β1 induced Smad2 expression, and activation were suppressed by Bcl11b overexpression (Fig. 4C, E, and F). Surprisingly, the levels of Smad3 and pSmad3 had no obvious change in either group (Fig. 4C, E, and F). These results indicated that overexpression of Bcl11b might restrain IL-17A expression through suppressing the TGFβ/Smad2 signaling pathway.
DISCUSSION
It is widely recognized that CD4+ T cells play an essential role in adaptive immune responses in allergic asthma. Based on specific cytokines present at the time of naïve T cell activation, CD4+ T cells differentiate into various effector T cell subsets including Th1, Th2, Th9, Th17, and Treg. Although the imbalance of Th1/Th2 reaction is a major pathological phenomenon of asthma,34 studies have shown that half of asthmatics are characterized by non-IgE-dependent and noneosinophilic inflammation.19 Recent evidence suggests that Th17 cells significantly enhance both neutrophilic and eosinophilic inflammation in asthma.52021 Specifically, severe and fatal asthma is considered to be associated with neutrophil-mediated disease rather than classical Th2-driven eosinophilic asthma.67
Polarization to the Th17 subset generally requires the presence of TGF-β and proinflammatory cytokines IL-6, IL-1β, and IL-23, resulting in the activation of transcriptional regulator RORγt (RAR-related orphan receptor γt) and RORα.2223 Th17 cells secret IL-17A, IL-17F, IL-21, and IL-22, which can aggrandize airway reactivity, allergic inflammation, airway remodeling, mucus cell metaplasia and smooth muscle hyperplasia in asthma.2425 However, the detailed molecule mechanism controlling IL-17A expression in asthma has not yet been clearly elucidated. In the current study, we detected decreased Bcl11b but increased IL-17A expression in patients with asthma, and revealed a strong negative correlation between Bcl11b and IL-17A in severe asthmatics. We also observed decreased Bcl11b expression and increased IL-17A production in the HDM-induced allergic airway inflammation models accompanied by higher levels of AHR, inflammatory cell infiltration, HDM-specific IgE and IgG1, and Th2 cytokines. To the best of our knowledge, this is the first report to demonstrate an inverse relationship between Bcl11b and IL-17 expression in asthma.
Bcl11b is a zinc finger protein that can act as either a transcriptional repressor or an activator, depending on the histone-modifying complexes recruited to the promoter.2627 Bcl11b is indispensable for multiple developmental checkpoints for thymocyte development including DN2, DN3, DP and SP stages.9101128 In mature T cells, inhibition of Bcl11b may affect the clonal expansion and cytolytic activity of CD8+ T cells.12 Bcl11b is also critical to maintain the suppression function of Treg cells. Bcl11b−/− Treg cells express reduced Foxp3 and IL-10 levels and up-regulated proinflammatory cytokines including TNF-α, IFN-γ, and IL-17.13 In an experimental autoimmune encephalomyelitis (EAE) model, Bcl11b restricts Th2-lineage gene expression in Th17 cells by repressing Gata-3 expression.15 These data implicate that Bcl11b is an important regulator in T cell differentiation and cellular function. In addition, our previous study revealed that Bcl11b could affect the proliferative ability of Th cells and that there was aberrant expression of the TGF-β/Smad pathway after altering the Bcl11b expression in naïve T cells.16 Therefore, it is reasonable to speculate that Bcl11b might regulate the expression of IL-17 through the TGF-β/Smad signaling pathway.
Many studies demonstrated that TGF-β is the quintessential cytokine of Th17 cell differentiation. TGF-β binding to TGF-β receptor II on T cells leads to the recruitment of TGF-β receptor I to the complex, resulting in the activation of the cytoplasmic transcription factors Smad2 and Smad3, which are associated with a variety of nuclear factors regulating gene transcription.29 It has been known that Smad2 and Smad3 must be phosphorylated to be active and then translocate to the nucleus in a complex with Smad4 to regulate target gene transcription.30 Our study provides in vivo evidence for the involvement of the TGF-β/Smad pathway in the HDM-induced asthma model with significantly up-regulated TGF-β signaling components, TGF-β1 and pSmad2/3. The inverse correlation between Bcl11b and TGF-β1/IL-17 suggests a possibility that Bcl11b negatively regulates TGF-β1 and/or TGF-β/Smad signaling to suppress IL-17A expression in asthma.
In vitro transfection of HDM-stimulated splenocytes with Bcl11b suppressed IL-17A expression and Smad2 phosphorylation even in the presence of exogenous TGF-β1, suggesting that Bcl11b directly suppresses TGF-β/Smad signaling. Smad3 phosphorylation remained unchanged in cultured splenocytes, regardless of TGF-β1 treatment and Bcl11b overexpression, although increased Smad2 and Smad3 phosphorylation was seen in CD4+ T cells purified from mice with HDM-induced allergic airway inflammation. These results demonstrated that Bcl11b contributes to the inhibition of IL-17 expression and IL-17-mediated inflammation through selective restraint of Smad2 activation rather than Smad3 activation.
In conclusion, results reported in this study demonstrate that Bc111b is a negative regulator for IL-17 expression in HDM-induced asthma and identify a novel mechanism for Bc111b negatively regulating IL-17 expression via the TGF-β/Smad2 signaling pathway. These findings provide a better understanding of the molecular mechanism of Bc111b that might lead to new therapeutic targets for SA.
 XML Download
XML Download