

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Taurine (2-aminoethanesulfonic acid), a conditionally essential nutrient, is synthesized from amino acid cysteine or methionine and ubiquitously presented in animal tissues. High concentrations of taurine are found in heart, muscles, brain and eyes of mammals, and sea foods such as squids, clams, oysters are rich in taurine. Taurine has sulfur containing amino acid like structure and vitamin like biological functions such as heart rhythm, blood pressure, platelet aggregation, neural excitation, sperm motility, eye sight and bile acid synthesis (Huxtable, 1992). Taurine is known to be effective in protecting against cardiovascular diseases by affecting cholesterol metabolism, protecting blood vessels and blood cells from oxidative damages and controlling osmoregulation through membrane transport. Action mechanisms of taurine are not clear except its clearing action of cholesterol for bile acid synthesis in which taurine is used for conjugation to form the primary bile acid. Taurine, an organic osmolyte, regulates changes of cell volume and intracellular ion concentrations by controlling membrane ion permeability. Osmolarity-sensitive taurine efflux following cell swelling caused an increase in Na and K permeability and an anion channel blocker diminished this effect (Hall et al., 1996). Cellular accumulation of organic substances such as inositol, taurine and amino acids caused an opening of K and/or anion exit channels, and the extruded K+ is pumped in by Na-K ATPase (Friedrich et al., 2006). Na-K ATPase acts on Na reabsorption in distal tubule and collecting duct of kidney cell. Na-K ATPase is reduced in patients with insulin resistant diabetes, obesity and hypertension (Hakam et al., 2006; Rabini et al., 1999). Angiotensin receptor agonists inhibit proximal tubule Na-K ATPase in obese hypertensive rat and Na induced hypertension was accelerated with taurine deficiency (Hakam et al., 2006). Platelet activating factor (PAF) released during certain pathological conditions such as ischemia may act on Na-K ATPase. PAF inhibited Na-K ATPase in rat cerebral cortex and these effects were abolished by a PAF antagonist. Stimulation of PAF receptor decreased Na-K ATPase in rat proximal tubule contributing to natriuresis (Handa et al., 2003). In in vitro study of platelet incubated with LDL, IDDM LDL significantly increased platelet aggregation and significantly reduced platelet membrane Na-K ATPase compared with control LDL (Rabini et al., 1999).
The purpose of this study was to examine the effect of taurine on CVD related three parameters using Sprague Dawley rats fed cholesterol diet. We measured plasma and liver levels of cholesterol, erythrocyte Na efflux through Na-K ATPase and Na passive leak and platelet aggregation using Sprague Dawley rats fed cholesterol diet.
Materials and Methods
Animals and diets
Forty of four weeks old Sprague Dawley rats (Orient Co Ltd, Gapyung, Korea were divided into four groups and fed ad libitum diets containing 0.5% cholesterol plus 0.2% cholate and/or 5% taurine along with the control diet for four weeks (Table 1). Rats had free access to water and were housed individual cages in a room maintained at 20-25℃ with a 12-hour dark-light cycle. Blood samples were obtained by cardiac puncture into heparinized vacuum tubes, and platelet aggregation and erythrocyte Na efflux were performed with fresh blood. Liver samples were prepared for microscopic examination and plasma and liver samples were stored at -70℃ for later assays.
Plasma and liver lipid assays
Liver lipids were extracted by a modified Folch method (Folch et al., 1957). One gram of liver tissue was homogenized in 6 ml of Folch solution [chloroform(2):methanol(1)] and 2 ml H2O for 5 min and centrifuged. The lower phase contained liver lipids. Lower phase of lipid fractions after being treated with triton X-100:chloroform (25 µl:475 µl) for total cholesterol and methanol for triglyceride was assayed using enzymatic kits (Asan Pharmaceuticals, Korea).
Erythrocyte Na efflux
Na efflux from erythrocyte occurs by at least three different pathways; Na-K ATPase, Na-K cotransport and Na passive leak. Na-K ATPase can be defined as the ouabain sensitive Na efflux through the pump and can be measured from the difference of the efflux rates into MgCl2 medium with and without ouabain. Na leak is defined as the Na efflux through passive Na channel occurring under inhibition of ouabain sensitive Na-pump and furosemide sensitive Na+-K+ cotransport and can be measured from Na efflux into a choline chloride medium containing ouabain and furosemide (Garay et al., 1979; Smith et al., 1984).
Red cell preparation: Blood was centrifuged at 1,000 × g for 10 minutes, and the plasma and buffy coat were removed. Red blood cells were washed 5 times with a cold isotonic washing solution [150 mM choline chloride, 10 mM Tris-4 morpholinopropane sulfonic acid (MOPS), pH 7.4 at 4℃], centrifuging at 1,000 × g for 5 minutes after each wash. The RBC pellet was resuspended in the choline chloride washing solution to give 40-50% hematocrit. The hematocrit was measured. A 50 µl aliquot of the RBC suspension was added to 5 ml of 0.025% acationox (a metal free detergent, Scientific Products, McGaw Park Illinois, USA to be used for determination of intracellular Na concentrations.
Na efflux: Four ml each of the RBC suspension were added to 40 ml MgCl2 medium with or without ouabain (70 mM MgCl2, 10 mM KCl, 85 mM sucrose, 10 mM glucose, 10 mM Tris-MOPS, pH 7.4 at 37℃, 1 mM ouabain). Two ml of the RBC suspension was added to 40 ml of choline chloride medium (150 mM choline chloride, 10 mM glucose, 1 mM ouabain, 1 mM furosemide, 10 mM Tris-MOPS pH 7.4 at 37℃). The RBCs in each medium were mixed and aliquoted in each of 12 tubes. Tubes in duplicate were transferred to an ice bath after incubation in 37℃ shaking water bath for 0, 2, 4, 6, 8 and 10 min for Na efflux through the pump and 0, 10, 20, 30, 40 and 50 min for the passive leak of Na. Tubes were centrifuged at 1,000 × g for 5 min, then the supernatant was removed and measured for Na concentration using Atomic Absorption Spectrophotometer (Shimadzu model AA6701F).
Calculations (Kang & Smith, 1990);
Na µg/(ml × min) × 60 min × mmole/23 mg × [44 ml-(4 ml × hct)/(0.0041 × hct)] × (mg/1000 µg) = mmole/ℓ rbc/hour (Na efflux)
Na µg/(ml × min) × mmole/23 mg × 101/hct × (mg/1000 µg) = mmole/ℓ rbc (intracelluar Na)
Platelet aggregation
Platelet aggregation was measured using a Chronolog Whole Blood Aggregometor (model 500-Ca, Havertown, Pennsylvania, USA). The whole blood was diluted with isotonic saline (1:2) to give platelet concentration 400,000/µl. Adenosine diphoshate (ADP, 2 µM) was added to initiate aggregation, and three readings of impedance changes were averaged for each rat. Platelet aggregation caused an increase in impedance(Ω) across two platinum electrodes. The impedance method using the fresh whole blood has the advantage of measuring platelet aggregation under nearly physiological conditions in the presence of other blood components.
Histological examination
Two parts of liver samples each rat were obtained from 4 rats each group. After being washed with saline, samples were fixed in 10% formalin for 48 hours and dehydrated in xylene for 18 hours. Samples were then embedded in paraffin and cut into 4-5 um cross section. Tissues were stained with hematoxylin-eosin for microscopic observation.
Results
Weight gain, food efficiency and liver/body weight ratio
Weight gain, food efficiency and liver/body weight ratio are listed in Table 2. Average daily gain (ADG) of taurine group was smallest among the groups with the lowest final weight, but not statistically different. Food efficiency ratio (FER, weight gain/food intake ratio) of taurine group was significantly decreased compared to other groups (p<0.05). Group fed taurine plus cholesterol recovered food intake and daily weight gain increasing the final weight to the control level. Liver/body weight ratio was significantly increased in groups of cholesterol and cholesterol plus taurine compared to the group of control or taurine (p<0.01). Taurine did not reduce the liver weight increased by feeding cholesterol diet.
Plasma and liver cholesterol and triglyceride
Plasma total cholesterol was significantly increased in groups fed cholesterol or cholesterol plus taurine compared to groups of control or taurine (p<0.05). Plasma total cholesterol of taurine group was lower than that of the control, but not significantly different (Table 3). However, the group fed cholesterol plus taurine had significantly lower plasma total cholesterol than the group fed cholesterol (p<0.05). Cholesterol lowering effect of taurine was more intensive in the cholesterol fed state. HDL-cholesterol was decreased in groups fed the cholesterol diet regardless of taurine supplementation and the difference between groups with or without cholesterol was significantly different (p<0.01).
Erythrocyte Na efflux
Intracellular Na tended to be lower in group fed cholesterol or taurine compared to the control group and higher in group fed cholesterol plus taurine compared to the control group (Table 4). Na efflux through Na-K ATPase and Na passive leak was somewhat reduced in group fed cholesterol or taurine and was augmented in group fed cholesterol plus taurine compared to the control, which showed a similar trend to the intracellular Na. Taurine supplementation caused a suppression of Na efflux in group fed control diet and restored the suppressed Na efflux in group fed cholesterol.
Platelet aggregation
As listed in Table 5, platelet aggregation was significantly decreased in the group fed taurine compared to the control (p<0.05) and the group fed cholesterol plus taurine was also lower in aggregation than the group fed cholesterol. Aggregation with high initial slope usually has high maximum aggregation as shown in the present study. Aggregation with high initial slope also showed a sharp dissociation curve which may be associated with platelet age and survivability.
Discussion
Tufft et al. (1992) reported that dietary taurine did not affect weight gain but improved feed efficiency during the first week of the experiment in broiler cockerels. However, taurine decreased their overall feed efficiency. On the other hand, taurine depletion caused a significant growth depression before retinal degeneration in monkey (Hayes et al., 1980). Dietary taurine suppressed weight gain with reduced food intake in Wistar rats, but not in Sprague Dawley rats when they were fed high cholesterol diet (Yamamoto et al., 2000). In the present study, taurine deceased weight gain and food efficiency, but taurine with cholesterol recovered the reduced weight with increased food intake. Huxtable et al. (1976) suggested earlier that taurine might have a suppression effect on eating and drinking by specially depressing the area of hypothalamus for stimulating eating and drinking. Taurine reduced Ca dependent GABA release in mouse cerebral cortex and GABA is known to play a role as potent stimulus for feeding through orexin mechanism in appetite control (Burdakov et al., 2003; Kontro et al., 1987).
Hypocholestrolemic action of taurine can be speculated on the activation of cholesterol 7α-hydroxylase, suppression of HMG-CoA reductase, ileal bile acid reabsorption and perhaps LDL receptor. Taurine supplementation induced the gene expression of cholesterol 7α-hydroxylase in diabetic rats fed high cholesterol diet, enhancing cholesterol degradation (Mochizuki et al., 1999). Murakami et al. (2002) reported that hepatic activity of cholesterol 7α-hydroxylase was doubled with taurine and taurine decreased cholesteryl ester contents, accompanied by decrease in acyl CoA:cholesterol acyltransferase (ACAT) activity. Taurocholate is a potent down regulator of both HMG CoA reductase, the rate limiting enzyme for cholesterol synthesis, and cholesterol 7α-hydroxylase, the rate limiting enzyme for bile acid synthesis (Pandak et al., 1992). Shefer et al. (1992) observed that cholesterol 7α-hydroxylase activity was increased with cholestyramine and cholesterol feeding and decreased with taurocholate in rats. Both cholesterol and taurocholate administration suppressed total microsomal HMG-CoA reductase activity by 87% and 65%, respectively. Taurocholate may be a far more potent inhibitor of cholesterol 7α-hydroxylase than other bile acids through feedback inhibition mechanism on bile acid synthesis (Shefer et al., 1992). Up-regulation of the LDL receptor and subsequent increase in receptor-mediated LDL turnover can be another cholesterol-lowering event in hamsters fed taurine. (Murakami et al., 2002). Chen et al. (2004) observed that taurine did not affect LDL receptor protein level in C57BL/6 mice and the decrease in cholesterol in the circulation was mainly due to its suppressive effect on the concomitantly secreted TG from the liver. Cholesterol feeding caused a remarkable increase in liver triglyceride and total cholesterol (Chen et al., 2004; Liu et al., 1995; Sobolova et al., 2006). Taurine supplementation resulted in markedly reduced hepatic secretion of cholesterol ester, preventing the hepatic accumulation of cholesterol in cholesterol fed rats (Chen et al., 2004). The present study showed that taurine supplementation caused decreased plasma and liver total cholesterol in both control and cholesterol fed groups. High cholesterol diet has been involved in increased total cholesterol and decreased HDL-cholesterol in different species (Oh et al., 2002; Sobolová et al., 2006; Sorci-Thomas et al., 1989). Sorci-Thomas et al. (1989) suggested that accelerated apo A-1 clearance from plasma may contribute to the reduction of HDL concentration resulting from cholesterol enriched diets. Cholesterol diet resulted in the accumulation of liver triglyceride with decreased plasma triglyceride (Liu et al., 1995; Oh et al., 2002), which can be explained with increased de novo synthesis of liver triglyceride and decreased secretion of triglyceride to the blood (Liu et al., 1995). Hypolipidemic effect of taurine was far greater in rats fed control diet than those fed cholesterol diet.
Cholesterol has been reported to be involved in membrane transport of cations. In vitro cholesterol enrichment of human erythrocyte caused an enzymatic inhibition of Na-K ATPase and decreased Na Efflux (Lijnen et al., 1995). Kurup et al. (2003) reported that plasma HMG CoA reductase activity was inversely correlated with erythrocyte membrane Na K ATPase in Alzheimer patients. Tranquilli et al. (2004) observed that higher cholesterol/phospholipid ratio was correlated with the reduced erythrocyte Na-K ATPase in pregnant women. Lower membrane fluidity with high plasma cholesterol/phospholipid ratio may be associated with lower Na-K ATPase which caused the progression of hypertension in pregnant women (Tranquilli et al., 2004). Mayol et al., (1999) reported that cholesterol enrichment significantly reduced Na-K ATPase in cultured human endothelial cells. The present study showed that cholesterol supplementation reduced erythrocyte Na-K ATPase as in previous studies. Taurine in human erythrocyte activated ouabain sensitive Na-K ATPase in the presence of calcium (Igisu et al., 1976). Qi et al. (1995) reported that taurine restored the depleted erythrocyte membrane Na-K ATPase activity due to cholesterol enrichment in in vitro system. In the present study, taurine supplementation rather reduced erythrocyte Na-K ATPase, but taurine with cholesterol tended to increase Na-K ATPase. Mrsny et al. (1985) reported that taurine and hypotaurine reduced hamster sperm cell Na-K ATPase for sustaining sperm motility. Taurine on Na-K ATPase may function differently depending upon cell types and numerous methods such as phosphate production, substrate concentration and Na efflux can be used as parameters for Na-K ATPase activity. Na leak is Na passive transport driven by intracelluar concentration of Na. Na efflux through Na leak in the present study was matched with the intracellular Na. High intracellular Na may be associated with decreased Na-K ATPase activity, followed by increased intracellular fluid (Lijnen et al., 1995). Zicha et al. (2001) reported an opposing result that abnormally high erythrocyte Na-K ATPase was a consequence of elevated erythrocyte Na content and associated with abnormal cholesterol metabolism in Dahl salt sensitive rats. In the present study, high Na-K ATPase with high intracellular Na in control or taurine plus cholesterol is rather consistent with the results of Zicha. Taurine and β-alanine are known to be cotransported into cell with Na ions and then Na ions are pumped back out against concentration via Na-K ATPase. More studies should be done to clear the action mechanism of taurine and cholesterol on Na-K ATPase and intracellular volume.
Platelet from hypercholesterolemic patient was more reactive to platelet activating factor (PAF) and the maximal aggregation was correlated with plasma PAF acetylhydrolase and total cholesterol (Elisaf et al., 1999). Platelet from hypercholesterolemic rat was more sensitive to thrombin and showed higher aggregability, and thrombin generation in the 2nd phase of platelet aggregation was far higher in hypercholesterolemic rats than the control (Winocour et al., 1990). In the present study, platelet from cholesterol fed rats was less sensitive in ADP induced whole blood platelet aggregation compared to those from control rats. However, taurine caused decreased platelet aggregation with or without cholesterol supplementation. Taurine has been considered as a modulator of platelet function. Taurine functions on platelet aggregation through PDGF-BB receptor dephosphorylation in vascular smooth muscle taurine, causing a significant suppression of PDGF-induced endothelial cell proliferation (Yoshimura, 2005). Taurine suppressed calcium induced platelet aggregation in vivo and ex vivo in vegetarians (McCarty, 2004). Taurine functions physiologically to dampen the calcium influx evoked by aggregating agonists, thereby down-regulating platelet aggregation. Platelet taurine causes a decrease in sensitivity of platelet to aggregating agonists such as thrombin, ADP, epinephrine, collagen and arachidonate.
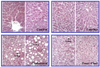
Livers from rats fed control and taurine were both normal and not different on gross observation. However, livers from rats fed cholesterol and cholesterol plus taurine were much larger and hypochromic, defined as fatty liver. On microscopic observation (Fig. 1), liver specimens from cholesterol plus taurine showed less fat drops than those from rats fed cholesterol, which suggests that taurine can lessen fatty liver in some extent when animal is on the hypercholesterolemic diet.





 XML Download
XML Download