

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Newcastle disease (ND) is an acute contagious avian disease caused by Newcastle disease virus (NDV) and has resulted in huge losses to the poultry industry [4]. ND has been listed as a notifiable terrestrial animal disease by the World Organisation for Animal Health (OIE) [18]. Currently, vaccination is the best choice to provide protection against NDV infection. The LaSota live vaccine strain is one of the NDV vaccine strains that are used widely in China. However, due to selection pressure resulting from vaccination, NDV variants continue to emerge, and ND outbreaks can still occur in vaccinated birds. The many defects in traditional vaccines for ND prevention encouraged us to develop a more effective vaccine for the prevention and control of ND infection.
Recent studies have shown that live attenuated Salmonella is an effective vector for delivery of heterologous antigens or DNA to the immune system [17182337]. In addition, live vectors can mimic the natural process of Salmonella infection through the oral route, resulting in mucosal and systemic immune responses against the pathogen's antigen [7]. In addition, live vectors can induce immune responses to aid in resisting the Salmonella infection. NDV is a member of the Paramyxoviridae, and its envelope contains two glycoproteins: virus hemagglutinin-neuraminidase (HN) protein and fusion protein (F). Both HN and F are important proteins for virus infectivity and pathogenicity [25]. Moreover, they can induce virus neutralizing antibody responses and act as protective antigens of NDV [510]. It has been reported that the F protein of NDV strain JS5 can be expressed in Vero cells by using attenuated Salmonella enterica serovar Typhimurium ZJ111 strain as a transgenic carrier [11]. In addition, the recombinant-attenuated Salmonella enterica serovar Typhimurium strain SL7207 harboring the F gene of NDV strain JS5 can induce preferable immune responses in chickens [28]. However, no reports have shown whether the HN protein of NDV strains can be expressed through an attenuated Salmonella strain to produce a corresponding immune response.
Salmonella Pullorum is currently a major bacterium that produces significant damage to the poultry industry. It is easy to bring forth biological safety problems by drug/vaccination over- or misuse, and, until now, there was no effective vaccine to protect against S. Pullorum. Attenuated S. Pullorum could be used as a novel live vaccine to prevent ND in chickens; however, it is unknown whether attenuated S. Pullorum could be an effective delivery system for the NDV HN gene, or whether it could be effective against challenges of NDV virulent viruses and S. Pullorum virulent strains. Therefore, in the present study, we explored the immunobiological characteristics of a recombinant S. Pullorum strain that included the major protective HN gene of NDV; the HN gene was carried by an asd mutant balanced-lethal system in the basic gene deletion-induced attenuated C79-13Δcrp strain.
Materials and Methods
Strains, plasmids, and viruses
The live attenuated S. Pullorum strain, C79-13ΔcrpΔasd, was a derivative of S. Pullorum C79-13 incorporating a deletion of the crp gene in the aromatic amino acid biosynthetic pathway and the asd gene. The strain was constructed and conserved by our laboratory. The prokaryotic expression vector pYA3493 (asd+, pBR322 ori, derivative β-lactamase signal sequence) and its host χ 6097 (F− ara Δ(pro-lac) rpsL ΔasdA4 Δ[zhf-2::Tn10] thi φ80dlacZΔM15) were kindly provided by Dr. Roy Curtiss III, University of Washington, USA. The asd attenuated strains were grown in LB broth containing 50 µg/mL diaminopimelic acid at 37℃. All bacterial strains were stored at −80℃ in growth medium containing 15% glycerol. The NDV virulent strain F48E9 and the vaccine strain LaSota were originally obtained from the China Veterinary Culture Collection. The NDV live vaccine LaSota was obtained from a commercial manufacturer (Liaoning Yikang Biological, China).
Construction of the live attenuated C79-13ΔcrpΔasd (pYA-HN) strain
Vector pYA3493 harboring the asd gene was a balanced-lethal host-vector system which was used to complement the chromosomal asd mutant strain C79-13ΔcrpΔasd. Specific primers for amplifying the NDV HN fragment were designed and synthesized based on the viral genome sequence of the vaccine strain LaSota (GenBank accession No. AF077761). Forward primer pH1: 5′CGGGATCCATGGACCGCGCCGTTAGC3′; reverse primer pH2: 5′CCCAAGCTTCTAGCCAGACCTGGCTTC3′ contained the BamH I and Hind III restriction enzyme sites, respectively, which are underlined at the 5′end. The HN fragment was amplified from the cDNA of the LaSota virus strain and was obtained following standard polymerase chain reaction (PCR) conditions using primer pairs pH1 and pH2. The PCR product and the plasmid vector pYA3493 were digested with BamH I and Hind III, respectively, and were purified by using a gel extraction kit (Axygen Scientific, USA). Subsequently, the HN gene was ligated to the plasmid pYA3493 using a Ligation Kit (Takara Bio, Japan) and transformed into Escherichia coli χ6097. The clones were confirmed by PCR amplification, restriction enzyme digestion, and gene sequencing. Finally, the pYA3493 plasmid and the identified recombinant pYA3493-HN plasmid were transformed into S. Pullorum C79-13ΔcrpΔasd by electroporation as previously described [31]. The asd+ transformants were selected on LB plates, and the final positive strain, named C79-ΔcrpΔasd (pYA-HN), was used as a potential live NDV vaccine candidate for the following oral immunization.
Expression of NDV HN proteins in S. Pullorum C79-13ΔcrpΔasd
To confirm the expression of HN protein from C79-13ΔcrpΔasd (pYA-HN), western blot analysis was performed as previously described [34]. Briefly, strains C79-13ΔcrpΔasd (pYA) and C79-13ΔcrpΔasd (pYA-HN) were cultured in LB broth at 37℃ to an optical density at 600 nm (OD600) of 0.8. Then, the total cells were harvested and broken ultrasonically. Protein samples were resuspended in 2× sodium dodecyl sulfate (SDS) sample buffer. The protein samples were then separated by performing 15% SDS-polyacrylamide gel electrophoresis after being boiled at 95℃ for 5 min. Separated proteins were stained with Coomassie blue and transferred onto nitrocellulose membranes. After overnight blocking (5% skim milk with 0.01% Tween 20 in phosphate buffered saline [PBS]) at 4℃, membranes were incubated with polyclonal antibody against NDV HN protein and alkaline phosphatase (AP)-conjugated goat anti-chicken IgG (Sigma-Aldrich, USA). Visualization of immunobinding was conducted by enhanced chemiluminescence (ECL) by using an ECL Plus detection kit (Thermo Scientific, USA).
Experimental chickens and bacterial immunization
Experiments were conducted in specific-pathogen-free (SPF) White Leghorn chickens that had been obtained from Jinan SPF Egg & Poultry, China. All chickens were housed in separate isolators in the facility of Laboratory Animal Care of Henan University of Science and Technology. Animal management and experimental procedures were carried out following the requirements of the Animal Care and Ethics Committees of Henan University of Science and Technology.
Two hundred chickens were randomly divided into four groups (n = 50 per group). The four groups were orally immunized with C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), or PBS by using oral gavage; the NDV live vaccine LaSota was used as the positive control (administered via intraocular-nasal vaccination with 106 EID50/chicken; EID50, 50% egg infective dose) at 7 days of age. Briefly, C79-13ΔcrpΔasd (pYA-HN) and C79-13ΔcrpΔasd (pYA) were cultured at 37℃ in LB broth supplemented with diaminopimelic acid (50 µg/mL) and shaking for 18 h. Freshly cultured bacteria were harvested, washed twice with PBS, and resuspended in PBS; next, each chicken was orally inoculated with 0.2 mL of bacterial suspension containing 1 × 109 colony-forming units [32].
Safety examination of the C79-13ΔcrpΔasd (pYA-HN) strain
To evaluate the safety of the C79-13ΔcrpΔasd (pYA-HN) strain, the general condition of each group of chickens was observed every day for 4 weeks after immunization. The typical clinical signs of infection (such as anorexia, depression, and diarrhea) and mortalities of S. Pullorum C79-13ΔcrpΔasd (pYA-HN) chickens were recorded regularly. In addition, fecal samples of three immunized chickens were collected and cultured; confirmation of C79-13ΔcrpΔasd (pYA-HN) strain presence was performed by undertaking PCR using SP-specific and HN-specific primers, as previously described, at 7, 14, and 21 days post-immunization (dpi). The C79-13ΔcrpΔasd (pYA) strain was used as a parallel control group in this study [26].
Body and organ weight evaluation
In order to evaluate the effects of the C79-13ΔcrpΔasd (pYA-HN) strain on growth rate and lymphoid organ weight, six chickens were randomly selected from each group and their body weights determined at 7, 14, 21, 28, and 35 dpi. Then, the six birds from each group were sacrificed and their lymphoid organs (spleen, bursa, and thymus) collected and weighed. Lymphoid organ to body weight ratios were calculated according to the following formula: (lymphoid organ weight [g]/body weight [g] × 100) [322].
Hemagglutination inhibition (HI) antibody titers examination by HI assay
In order to assess the HI antibody titers, HI assays were carried out according to a standard protocol previously described [13]. Briefly, serial two-fold dilutions of each serum sample with PBS were prepared in 96-well plates; then, an equal volume of four hemagglutination units. of NDV were added to each well and mixed. The mixture was incubated for 20 min at room temperature. Finally, a 1% SPF chicken erythrocyte suspension was added to each well and hemagglutination was allowed to proceed for 40 min at room temperature. The highest dilution of serum that completely prevented hemagglutination was scored as the HI-titer and was expressed in log2 scale.
Serum immunoglobulin G (IgG) and intestinal secretory IgA (sIgA) examination by enzyme-linked immunosorbent assay (ELISA)
Anti-S. Pullorum specific antibodies were examined by indirect ELISA as previously described. Briefly, serum samples were collected from six chickens of each group on 7, 21, 28, and 35 dpi. The 96-well ELISA plates were coated with 10 µg/mL bacterial protein of S. Pullorum C79-13 which followed by overnight incubation at 4℃ and were then blocked with 100 µL/well 5% skim milk for 2 h at 37℃. Test serum samples at 1:500 dilution (optimum dilution determined by chessboard titration) of 100 µL/well were added to the plates and incubated overnight, followed by washing with 0.05% PBS-tween-20 and incubation with 1:10,000 HRP (100 µL/well) conjugates of rabbit anti-chicken IgG (GenScript, China) for 1 h. Finally, TMB substrate solution (100 µL/well) was added for color development. The reaction was stopped by using 2 mol/L H2SO4 100 µL/well. Absorbance was determined at 450 nm with an ELISA reader. Each serum sample was assayed in triplicate.
Intestinal sIgA level was determined according to a method previously described [2426]. Intestinal lavage samples were collected from six chickens of each group at 7, 14, 21, 28, and 35 dpi and stored at 4℃. The sIgA concentrations were quantified by using chicken sIgA ELISA quantitation kits (eBioscience, USA), according to the manufacturer's instructions and using a dilution of 1:4 of intestinal washes.
Lymphocyte proliferation assay
The lymphocyte proliferation assay was performed as previously described [32], but with some modifications. Briefly, 1 mL of blood collected from the wing vein and twice diluted with PBS was overlain on the same volume of lymphocyte separation medium (1.077 specific gravity; Sigma-Aldrich) and centrifuged at 500 × g for 20 min. The peripheral lymphocytes were harvested and checked by staining with trypan blue dye. Viable lymphocytes suspension (5 × 105 cell/well) were plated into 96-well tissue culture plates in triplicate and incubated in 100 µL complete RPMI-1640 medium with or without NDV (104 EID50/well) and with 10 µL/well conA (0.5 mg/mL; Tiangen, China,) as a positive control. After 72 h, 10 µL/well MTT (5 mg/mL; Tiangen) were added to each well and the plates were incubated for 4 h at 37℃. Then, 150 µL per well of dimethyl sulfoxide was added and the plates were further incubated for 10 min at 37℃. OD was measured with an ELISA reader (Perlon, China) at a wavelength of 570 nm. All samples were analyzed in triplicate. The stimulation index (SI) was calculated according to the formula: SI = mean OD of stimulated cells/mean OD of un-stimulated cells [32].
Challenge experiment
To evaluate whether the recombinant-attenuated S. Pullorum strain could induce protective immunity against ND and S. Pullorum, the chickens (n = 10) in each group were orally challenged through the oculonasal route with 106 EID50 of highly pathogenic NDV F48E9 strain or with lethal dose of 70% (LD70) of S. Pullorum C79-13 field strain at three weeks after immunization. The challenged chickens were observed daily for a period of 14 days, and mortality and clinical signs of ND and S. Pullorum infection were recorded. Chickens were considered protected if they showed no clinical signs and survived the challenge period.
Results
HN expression of S. Pullorum C79-13ΔcrpΔasd (pYA-HN)
The PCR amplification, restriction enzyme digestion, and gene sequencing results revealed that the HN gene was correctly cloned into vector pYA3493 and confirmed that the pYA-HN plasmid was transformed into S. Pullorum strain C79-13ΔcrpΔasd. A protein band corresponding to 63 kDa was detected in C79-13ΔcrpΔasd(pYA-HN) by SDS-PAGE (panel A in Fig. 1). In order to confirm the protein band in 63 kDa was the HN protein, immunoblot analysis was conducted with anti-HN antibodies. The 63 kDa band, which was identical with the predicted size of the HN protein, was specifically recognized by the anti-HN antibody (panel B in Fig. 1).
Safety evaluation of the C79-13ΔcrpΔasd (pYA-HN) strain from fecal shedding
In order to validate the safety of the C79-13ΔcrpΔasd (pYA-HN) strain, chickens were observed for changes in their general condition for four weeks post-immunization. During that period, all chickens were visually healthy and did not show any typical signs of S. Pullorum disease (PD). After oral immunization with the C79-13ΔcrpΔasd (pYA-HN) and C79-13ΔcrpΔasd (pYA) strains, and after applying direct and enrichment culture methods at 7, 14, and 21 dpi, neither strain was isolated from fecal samples of the immunized groups.
Body and organ weights evaluation
As demonstrated in panel A in Fig. 2, compared to the PBS control group, the growth rates in C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), and LaSota-treated birds were not significantly different over the entire experimental period (p > 0.05). There was also no difference in spleen/body, thymus/body, or bursa/body weight ratios of birds among the groups immunized with C79-13 ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), LaSota, or PBS at 7 dpi and 14 dpi. However, at 21, 28, and 35 dpi, the spleen/body, thymus/body, and bursa/body weight ratios of birds immunized with C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), or LaSota were significantly higher (p < 0.05) than that of the PBS control birds. There was no difference between the C79-1ΔcrpΔasd (pYA-HN) strain and the C79-13ΔcrpΔasd (pYA) strain or the LaSota vaccine (p > 0.05) (panels B-D in Fig. 2).
Assessment of HI titer
To determine the HI antibody titer, serum samples of the experimental chickens were collected on days 0, 7, 14, 21, 28, and 35 after immunization. Post-immunization, the serum HI antibody titers were significantly induced in chickens immunized with the C79-13ΔcrpΔasd (pYA-HN) strain or the NDV live vaccine LaSota. The HI antibody titers were maximal at 21 days after immunization, after which the HI antibody titer declined continuously. The HI antibody titers in chicken immunized with C79-13ΔcrpΔasd (pYA-HN) were lower than those in chicken immunized with LaSota, but the difference was insignificant (p > 0.05) (Fig. 3). The results indicated that C79-13ΔcrpΔasd (pYA-HN) strain could effectively stimulate and increase HI antibody levels.
Assessment of serum IgG and intestinal sIgA by ELISA
To evaluate the specific humoral immune response, serum anti-S. Pullorum specific antibodies IgG and intestinal sIgA concentrations were determined at 7, 21, 28, and 35 days after immunization by performing ELISA. The mean OD values of serum IgG antibodies from the C79-13 ΔcrpΔasd (pYA-HN) strain and C79-13ΔcrpΔasd (pYA) strain immunized groups were significantly higher than those in the LaSota live vaccine-immunized group and the PBS control group (p < 0.01); however, there was no significant difference between the C79-13ΔcrpΔasd (pYA-HN) strain and C79-13ΔcrpΔasd (pYA) strain immunized groups (p > 0.05) (panel A in Fig. 4). Compared to the control group, the intestinal sIgA in the C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), and LaSota live vaccine groups observed 14 days after immunization increased consistently and reached the highest levels at the third week after immunization. The LaSota live vaccine group had the highest sIgA level, followed by the C79-13ΔcrpΔasd (pYA-HN) strain and the C79-13ΔcrpΔasd (pYA) strain (panel B in Fig. 4). These data suggested that the C79-13ΔcrpΔasd (pYA-HN) strain can stimulate a significant specific immune response.
Assessment of cell-mediated immune response by lymphocyte proliferation assay
To determine further the effect of the C79-13ΔcrpΔasd (pYA-HN) strain on cell-mediated immune responses, the lymphocyte proliferation assay was performed using the specific NDV antigen. The SI values of the chickens immunized with C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), LaSota live vaccine, or PBS were maximal at 21 days after immunization. The results revealed a significant (p < 0.05) increase in SI in chickens immunized with the C79-13ΔcrpΔasd (pYA-HN) strain and the LaSota live vaccine compared to those in the C79-13ΔcrpΔasd (pYA) and PBS groups; however, the SI of chickens immunized with the C79-13ΔcrpΔasd (pYA-HN) strain did not significantly increase over that in chickens immunized with LaSota live vaccine (p > 0.05) (Fig. 5).
Examination of protective efficacy against virulent NDV and S. Pullorum challenge
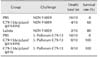
To determine whether the C79-13ΔcrpΔasd (pYA-HN) strain promotes protection against NDV and S. Pullorum, at 3 weeks after the vaccination the chickens were challenged with strong virulent NDV strain F48E9 at a dose of 106 EID50 in 0.2 mL or with the LD70 of S. Pullorum. Thirty days after viral challenge, the survival rates of chickens immunized with C79-13ΔcrpΔasd (pYA-HN) strain, LaSota live vaccine, and PBS were 60%, 80%, and 0%, respectively. Moreover, 30 days after the bacterial challenge, the survival rate of the chicken in C79-13ΔcrpΔasd (pYA), C79-13ΔcrpΔasd (pYA-HN) strain and PBS groups were 100%, 100%, and 0% (Table 1). The results indicated that the C79-13ΔcrpΔasd (pYA-HN) strain can promote specific immune protection against both NDV and S. Pullorum challenge.
Discussion
The use of attenuated Salmonella as a vector carrying foreign protein represents a novel approach to preventing and controlling infectious diseases of humans and animals. Although several new types of vaccine subspecies of NDV have been developed in past decades, there are several disadvantages of these vaccines in the control of ND. In this study, a recombinant-attenuated S. Pullorum strain secreting the NDV HN protein was constructed, and the expression of the HN protein from C79-13ΔcrpΔasd strains carrying the HN gene in the pYA3943 plasmid was demonstrated by western blot analysis (Fig. 1). The pYA3943 plasmid contains the pBR ori and asd genes, and it may be a balanced-lethal host-vector system to complement the chromosomal asd mutation strain; moreover, it can be stably maintained for over 50 generations in the mutational Salmonella [2038]. Therefore, the plasmid harboring the HN gene can be stably maintained in the attenuated S. Pullorum strain C79-13ΔcrpΔasd and can result in large amounts of HN protein being expressed.
The growth rates in all immunized groups were insignificantly different during the experimental period (p > 0.05); however, the spleen/body, thymus/body, and bursa/body weight ratios of the immunized birds were significantly higher (p < 0.05) than that of the PBS control birds at 21, 28, and 35 dpi (Fig. 1). These results indicate that the recombinant-attenuated S. Pullorum strain did not affect the growth rate in immunized birds, but it could effectively stimulate an increase in immune organ weight.
In the present study, in order to evaluate the level of humoral immune responses and cell-mediated immune responses of chickens that had been immunized with C79-13ΔcrpΔasd (pYA-HN), C79-13ΔcrpΔasd (pYA), or NDV live vaccine, the HI antibody titer and intestinal sIgA concentrations (detected for ND), serum anti-S. Pullorum specific antibodies IgG level (detected for PD), and the lymphocyte proliferation activity (detected for both ND and PD) were determined at different times after immunization. The sIgA antibody is the first line of defense against microbial infection, and it is the primary antibody mediating mucosal immunity [1415]. The mucosal immune effect of oral inoculations can be assessed by examining sIgA antibody levels, as well as by examining systemic antibodies and cell-mediated immune responses [1229]. Compared to the control group, the HI antibody titer, the intestinal sIgA levels of both C79-13ΔcrpΔasd (pYA-HN) strains and the NDV live vaccine-immunized groups all showed significant increases at the second week after immunization. The sIgA levels increased to the highest level at the third week after immunization; however, there was no significant difference between the C79-13ΔcrpΔasd (pYA-HN) strain and the NDV live vaccine groups (p > 0.05) (Figs. 3 and 4). The observed level of intestinal mucus sIgA showed that C79-13ΔcrpΔasd (pYA-HN) strain could colonize the intestinal lymph nodes and lead to an effective mucosal immune response. The anti-S. Pullorum specific serum antibodies IgG of both C79-13ΔcrpΔasd (pYA-HN) and C79-13ΔcrpΔasd (pYA) were significantly higher than those of the NDV live vaccine-immunized group and the PBS group, but the results from the C79-13ΔcrpΔasd (pYA-HN) and C79-13ΔcrpΔasd (pYA) immunized groups were similar (Fig. 4), showing that the existence of the HN gene did not affect the immune characteristics of the recombinant strains.
Cell-mediated immune responses can be represented by the SI of lymphocytes by using a specific antigen [219]. Significant cell-mediated immune responses were shown by the peripheral blood lymphocyte proliferation assay in both the C79-13ΔcrpΔasd (pYA-HN) and NDV live vaccine groups to be greater than that of the control group at 21 days after immunization (p < 0.05) (Fig. 5). These data suggest that the C79-13ΔcrpΔasd (pYA-HN) strain can effectively induce an increased level of acquired immunity, including mucosal immune responses.
Over the past 30 years, research into attenuated Salmonella as an oral live vaccine vector to carry and express exogenous protective antigen genes of pathogens has become popular. It has been reported that using attenuated Salmonella to construct recombinant live vaccines not only has the ability to activate the body's immune system to resist Salmonella infection, but it also can significantly enhance the immune effect of the exogenous gene being carried [1819273035]. In this study, the survival rates of the chicken challenged chicken with the C79-13 strain after immunization with C79-13ΔcrpΔasd (pYA-HN) or C79-13ΔcrpΔasd (pYA) were 100% (Table 1). These results suggest that the recombinant-attenuated S. Pullorum strain can induce a high protective effect against infection by Salmonella.
The NDV LaSota strain belongs to genotype II of NDV class II, and it has been used as an NDV live vaccine strain since the 1950s [616]. Several studies have reported that the LaSota vaccine can provide cross-protection against other NDV genotypes [93336]. In the present study, the recombinantattenuated S. Pullorum strain secreting the LaSota HN protein displayed 60% protective efficacy against challenge with the IX genotype NDV strain F48E9, with the LaSota live vaccine displaying 80% protective efficacy. Recently, Yang et al. [36] reported that the recombinant IBV H120 vaccine strain that expresses LaSota HN protein displayed 80% protective efficacy against NDV F48E9. Kumar et al. [21] reported that birds immunized with the recombinant avian paramyxovirus type 3 vector expressing HN proteins could partially protect against a virulent NDV strain; however, birds immunized with a recombinant avian paramyxovirus type 3 vector expressing NDV F protein alone or in combination with the recombinant avian paramyxovirus type 3 vector expressing HN proteins could completely protect against a virulent NDV strain. Therefore, the development of a recombinant-attenuated S. Pullorum strain that expresses both HN and F proteins needs to be further investigated to elucidate its potential protective efficacy against a virulent NDV. In our future studies, the recombinant strain will be used in combination with an ND classic vaccine strain (e.g., LaSota) in chicken, and the immunological protection effects and their mechanisms will be examined.
In conclusion, oral immunization with the recombinant-attenuated S. Pullorum C79-13ΔcrpΔasd(pYA-HN) strain can stimulate the production of the specific IgG antibody of S. Pullorum, the HI antibodies of NDV, and the intestinal mucosal immune antibody sIgA in chickens. In addition, the recombinant-attenuated S. Pullorum expressing HN could provide effective protection to chickens against the challenges of both NDV and S. Pullorum virulent strains. The results of this study suggest that the recombinant-attenuated S. Pullorum C79-13ΔcrpΔasd (pYA-HN) strain has the potential to be developed as a live attenuated bivalent oral vaccine candidate against both ND and PD.





 XML Download
XML Download