

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Protease inhibitors (PI) are a class of medications used to treat viral infections such as HIV (AIDS) and hepatitis C virus [2]. PI prevents viral replication by inhibiting the activity of protease, an enzyme used by the viruses to cleave nascent proteins for final assembly of new virons [2]. PI is a cocktail of water-soluble protease inhibitors with a broad specificity for the inhibition of serine, cysteine, and metalloproteases. It has been known that protease inhibitors act as antiretroviral, antiprotozoal and anticancer activity and they include 1) aminoethyle benzenesulfonyl fluoride and aprotonin which inhibit which trypsin, chymotrypsin, plasmin and thrombin, 2) EDTA- metalloproteases, 3) leupeptin hemisulfate salt as plasmin and cathepsin, 4) cystein proteases as leucin aminopeptidase and alanyl aminopeptidase.
The use of retroviral PI in conjunction with other antiretroviral drugs, has dramatically improved the morbidity and mortality associated with HIV infection [22]. However, patients under antiretroviral therapy develop a lipodystrophic syndrome as shown by altered body fat distribution and alterations of glucose and lipid metabolism [3,4,17,22]. In the blood, serum triglycerides and cholesterol are increased in these patients. There is also a high prevalence of hyperinsulinemia and insulin resistance, which results in diabetes [4,23]. It has been known that PI might inhibit adipocyte differentiation through the inhibition of peroxisome proliferators activated receptor gamma (PPARγ) and retonid X receptor that known to be a potent mediator of adipogenesis [1,21].
As known, fat deposition depends on accumulation of triacylglycerides in adipocytes [12]. Adipose tissue can convert glucose and amino acids into fatty acids when stimulated by insulin [10,14]. Adipose tissue can also secrete hormones and cytokines for its own regulation [13]. Human fat cells have been shown to have mRNA for adipsin (complement factor D), 3rd complement component (C3) and factor B, all of which are components necessary to generate activated C3 (C3a), the precursor for acylation stimulating protein (ASP). It was initially identified from human plasma [7], and determined to be a derivative of the C3. Arginine removal from C3a by carboxypeptidase results in C3a-desArg that is known as ASP [16].
Human and rodent adipocytes synthesis and secret complement factors such as C3, B and adipsin [5,8]. ASP acts on adipocyte membrane to increase glucose transport and diacylglycerol acyltransferase activity [11], so ASP influences tri-acyle-glycerol (TAG) biosynthesis in adipocytes [19]. Basal production of ASP in adipocytes is dependent on rate of adipocytes differentiation [6]. In addition, ASP stimulates glucose uptake through the translocation of glucose transporters Glut 1 and Glut 4 from the intracellular pool to the cell membrane [11]. Finally, similar to insulin, ASP inhibits hormone sensitive lipase (HSL) through stimulation of phosphodiesterase, thus inhibiting lipolysis in human adipocytes [20]. The relation between adipogenesis, PI and ASP is elucidated well. For that, we examined the effect of PI and ASP alone and in combination on adipocytes differentiation and triglycerides accumulation in 3T3-L1 cells.
Materials and Methods
Materials
Dulbecco's modified Eagles medium (DMEM), bovine serum albumin, bovine insulin and isobutylmethylxanthine (IBMX) were purchased from Sigma-Aldrich (USA). Fetal calf serum (FCS) was from Trace Scientific (Australia), dexamethasone, antibiotics (penicillin and streptomycin) and troglitazone were purchased from Wako Pure Chemicals (Japan). Protease inhibitor cocktail was from Sigma-Aldrich, USA and 3T3-L1 cells were from Zen-Bio, USA.
3T3-L1 cells differentiation and methodology
3T3-L1 cells were grown in DMEM containing 10% FCS and antibiotics (100 U/mL penicillin, and 100 µg/mL streptomycin). When cells reached confluence (referred to day 0), the cells were cultured in DMEM contains 0.5 mM IBMX, 1 µM dexamethasone, and 10 µg/mL insulin for 2 days, and subsequently in fresh DMEM containing 10% FCS, 10 µM troglitazone and 10 µg/mL insulin every 2 days for 12 days to achieve cell differentiation to mature adipocytes. To test the effect of PI on lipids accumulation in adipocytes, PI was added from day 4 to day 8 in doses of ×300, ×200, ×150 and ×100. To test the effect of PI and ASP, on day 4 of differentiation, the cells were incubated with insulin, PI alone or together for 4 days then stained with oil red O as shown down. To test the effect of ASP, on day 4 of differentiation, ASP in different doses different doses (Low, 16.7; Medium, 45 and High, 450 ng/mL) was added to insulin (10 µg/mL) alone or in combination with PI (×150) every day for 4 days and the cells were stained by oil red O.
RT-PCR analysis
Total RNA was extracted from culture adipocytes from day 0 (day of confluence) to day 12 of differentiation to test ASP expression.by the guanidine-isothiocyanate method using TRIzol reagent (Gibco, USA). RNA (1 µg) was treated at 72℃ for 5 min and reverse transcribed using 100 units of Moloney murine leukemia virus reverse transcriptase (Gibco, USA), 50 pmol of poly (dT) primer and 20 nmol of dNTPs in a total volume of 10 µL at 37℃ for 1 h. After heating at 94℃ for 5 min, PCR amplification was performed with 2.5 units Taq polymerase (Perkin-Elmer, USA), 3 mM MgCl2 and 50 pmol of forward and reverse primers specific for respective genes in a total volume of 50 µL. The primer pairs and PCR condition of ASP and glyceraldehydes-3-phosphate dehydrogenase (G3PDH) were as following: for ASP the primer pairs and PCR condition forward, 5'-GTCTTCACGGATGCAGGACTCA-3' reverse, 5'-T CAATGACGGTCCACAACC-3' the annealing temperature was 58℃ for 1 min to amplify a product of 460 bp. The reaction was carried out for 30 cycles. For bovine G3PDH, PCR condition was as follows, the annealing temperature was 59℃ for 30 sec, the forward primer: 5'-ACCACTGT CCACGCCATCAC-3' and the reverse primer: 5'-TCC ACCACCCTGTTTGCTGTA-3' the reaction was carried out for 25 cycles to amplify a product of 450 bp. After electrophoresis in 1.5% agarose gel, the PCR products were stained with ethidium bromide and visualized under UV lamp.
Oil red O staining
Cultured preadipocytes when reached confluence (day 0) and till 12 days of adipogenic differentiation were fixed with 10% formalin in isotonic phosphate buffer for 1 h, stained with 0.5% oil red O in 60% isopropyl alcohol for 1 h and rinsed extensively with water, lipid droplets were stained red. After visualization under binuclear ordinary microscope (×400), oil red O stained lipid droplets were extracted by 1 mL of absolute isopropyl alcohol and quantified by measuring absorbance in spectrophotometer at 540 nm.
Results
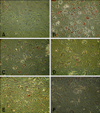
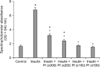
As seen in Fig. 1, the ASP expression was examined up to 12 days as seen it was time dependently increased using RT-PCR analysis. This increase in ASP expression was recorded in day 4 and reached the plateau at 8 days, and continued high up to 12 days. Next, the degree of lipids accumulation was examined microscopically in presence or absence of PI. The presence of insulin alone (10 µg/mL) increased the lipids accumulation. Addition of PI dose dependently inhibited adipogenesis and lipids accumulation (Fig. 2). When the degree of lipids accumulation measured specrophotometrically, the effect was clear and significantly (p < 0.05) increased with insulin (2 fold increase) and inhibited when co-treated with different doses of PI in comparing with insulin and control (Fig. 3).
To test that effect, cells were incubated with insulin and as seen in Fig. 4, the level of adipocytes differentiation and lipids accumulation was increased. Moreover, when it incubated with ASP in high dose (450 ng/mL) together with insulin, there was an additive increase in lipids accumulation was seen (Fig. 4). When the cells were incubated in presence of insulin and PI, TG accumulation was decreased. When ASP in low, medium and high doses of ASP was added together with insulin and PI, TG accumulation was partially reversed when compared with that of insulin and PI together (Figs. 4 and 5).
Discussion
The results presented in this study showed that PI inhibited TG accumulation and inhibition was partially reversed by addition of ASP. This study focused on the role of ASP in adipogenesis alone and in combination with PI, which were widely used as co-treatment therapy for hepatitis C and AIDS patients [2]. In this study, ASP expression was highly expressed during cell differentiation and maturation and confirmed the relation between ASP and TG accumulation; comparable finding was confirmed in another study in bovine adipose tissue explants [13].
Our data showed that ASP expression increased during differentiation of preadipocytes and same findings was reported in another study [15]. Furthermore, Koistinen et al. [15] reported that there was no increase in plasma ASP levels in response to an oral fat load. However, there was parallel correlation between the magnitude of postprandial lipemia and fasting ASP levels or subcutaneous adipose tissue C3 mRNA expression.
The exact mechanism of adipogenesis inhibition by PI is not well defined, but two pathways involved in that inhibition were confirmed, sterol regulatory element binding protein1 and PPARγ [2,19,21]. We can say that PI may inhibites ASP expression in mature adipocytes through modulation of other known genes that involved in TG biosynthesis as ASP and HSL [9,13]. In the study of De Barros et al. [9], the chronic lactacystin treatment of preadipocytes in an adipogenic medium led to a decrease of various differentiation markers such as triglyceride content, fatty acid binding protein-2 and HSL expression [9]. To our knowledge, this is the first investigation showing that PI cocktail has an inhibitory effect on 3T3-L1 preadipocytes differentiation is partially inhibited in presence of ASP.
It is known that PPARγ agonist are potent stimulators of adipocytes differentiation, so it is possible that PI induced its effect by inhibition of PPARγ expression, but Zhang et al. [25] found that PI failed to stimulate transcriptional activation of PPARγ in mammalian cells. Therefore, PI used for treatment of HIV inhibitors, may act as indirect inhibitors that antagonize the expression of adipogenesis related genes [2]. Xia et al. [24] reported that ASP is an important factor in regulating metabolic balance. The absence of ASP production results in increased energy expenditure in mice through its effect on TG synthesis and decrease peripheral energy expenditure [24]. ASP signaling involves sequential activation of PI3K, with downstream activation of protein kinase C, Akt and MAPK/ERK1/2, all of which lead to an effective and prolonged stimulation of TG synthesis [16]. In conclusion, this study showed the stimulatory effect of ASP and the inhibitory effect PI on adipogenesis in 3T3-L1 cells and that effect partially inhibited when cells co-treated with PI and ASP.



 XML Download
XML Download