

 PDF
PDF Citation
Citation Print
Print


Abbreviations
Ag85B
antigen 85B
AIM2
absent in melanoma 2
APC
antigen-presenting cell
BCG
Bacillus Calmette-Guérin
cGAMP
cyclic guanosine monophosphate-adenosine monophosphate
cGAS
cyclic guanosine monophosphate-adenosine monophosphate synthase
CTL
cytolytic T cell
DPR Korea
Democratic People's Republic of Korea
ESAT-6
early secretory antigenic target-6
GLA-SE
glucopyranosyl lipid adjuvant-stable emulsion
HIV
human immunodeficiency virus
IFN
interferon
IGRA
interferon-γ release assay
IRF3
interferon regulatory factor 3
MDR-TB
multi-drug resistant tuberculosis
MHC
major histocompatibility complex
MPL
3-O-desacyl-4′-monophosphoryl lipid A
NKT
natural killer T
OECD
Organization for Economic Cooperation and Development
PACTR
Pan African Clinical Trials Registry
STING
stimulator of interferon genes
TB
tuberculosis
TBK
TANK-binding kinase
Th1
type 1 T helper
TLR
Toll-like receptor
TNF
tumor necrosis factor
WHO
World Health Organization
α-GalCer
α-galactosylceramide
β2M
beta 2 microglobulin
INTRODUCTION
Tuberculosis (TB) is an infectious disease generally transmitted through aerosols containing Mycobacterium tuberculosis. TB has been responsible for the death of one billion people in the last 200 years (1) and is one of the 10 leading causes of death in the world (including people infected with human immunodeficiency virus [HIV]) (2). In 2015, approximately 10.4 million people worldwide had TB and 1.8 million people died of TB (2). Key risk factors for TB include poverty, overcrowding, malnutrition, alcohol abuse, and HIV infection (345). Most infections caused by M. tuberculosis are latent TB without symptoms; however, approximately 12% of these cases develop into active TB (26). Prior to 2001, the tuberculin skin test was the only diagnostic test for TB; however, in the 2000s, 2 interferon (IFN)-γ release assays (IGRAs) were approved by the Food and Drug Administration (FDA). In IGRA, the concentration of IFN-γ secreted by T lymphocytes in patients exposed to M. tuberculosis is measured after stimulation with early secretory antigenic target-6 (ESAT-6), which is the main antigen produced by M. tuberculosis (7). The first licensed IGRA was the QuantiFERON-TB Gold In-Tube test. This method measures the concentration of IFN-γ secreted into the peripheral blood of a patient after exposure to ESAT-6, CFP-10, and TB7.7. The second licensed IGRA was the T-SPOT.TB test, which is performed on whole blood. First, peripheral mononuclear cells are extracted from the collected blood sample and exposed to ESAT-6 and CFP-10, after which the concentration of secreted IFN-γ is measured. According to FDA-approved indications, diagnostic tests use different antigens; therefore, test results may be different. Consequently, it is recommended that these assays are considered indirect tests and are performed together with radiology or other medical evaluations (7).
Typical anti-TB drugs such as isoniazid, also known as isonicotinylhydrazide, and rifampicin are not effective against multi-drug resistant tuberculosis (MDR-TB) (8). Research on new anti-TB drugs is ongoing. Currently, bedaquiline (Sirturo®; Janssen Therapeutics, Titusville, NJ, USA) and delamanid (Deltyba®; Otsuka Pharmaceutical Co., Ltd., Tokushima, Japan) are being developed as treatments for MDR-TB (9). Other possible treatments include repurposed drugs such as linezolid, imatinib, and metformin (1011121314).
Currently, the only licensed vaccine for TB is the Bacillus Calmette-Guérin (BCG) vaccine, which is made by attenuating Mycobacterium bovis (15). Although vaccination with BCG prevents against tuberculous meningitis and disseminated TB in children, it does not prevent the development of a primary infection. Furthermore, it is not effective in preventing the reactivation of latent pulmonary infection. The efficacy of BCG vaccination is reported to vary between 0% and 80% for the prevention of pulmonary TB. In addition, it has limited efficacy in adults (1617). Therefore, the development of new TB vaccines is necessary. In this review, we have summarized the findings of studies on newly developed TB vaccines, the status of adjuvant research, and novel approaches for developing new TB vaccines.
EPIDEMIOLOGY
According to data from the World Health Organization (WHO), there were 10.4 million TB cases globally in 2015, which indicated an incidence of 142 cases per 100,000 people (2). The number of annual TB cases per country in 2015 was less than 10 per 100,000 in high-income countries and 150–300 per 100,000 in 30 countries considered as high-TB burden countries by WHO. Countries with an incidence rate of more than 500 per 100,000 people include Lesotho, Mozambique, and South Africa (2). The Republic of Korea ranks first in terms of TB incidence (80 persons per 100,000 people) and mortality (5.1 persons per 100,000 people) among the member countries of the Organization for Economic Cooperation and Development (OECD) (Fig. 1, Tables 1 and 2). These are above the OECD average TB incidence of 11.4 (per 100,000 population) and the average TB mortality of 1.0 (per 100,000 population) (Table 3). To solve this problem, the Korean government is conducting a national project to reduce TB incidence to 50 per 100,000 people by the year 2020.
Figure 1
Total TB cases and new TB cases from 2001 to 2016 in the Republic of Korea. The total cases and new cases of TB patients from 2001 to 2016 in the Republic of Korea are shown as the number per 100,000 individuals. The data are obtained from National Tuberculosis Management Guidelines (The Korea Centers for Disease Control and Prevention, 2017).

Table 1
Number of total TB cases and new TB cases in Republic of Korea (2007–2016)

Table 2
Number of deaths by TB in Republic of Korea (2006–2015)

| Year | Deaths |
|---|---|
| 2006 | 2,726 |
| 2007 | 2,376 |
| 2008 | 2,323 |
| 2009 | 2,292 |
| 2010 | 2,365 |
| 2011 | 2,364 |
| 2012 | 2,466 |
| 2013 | 2,230 |
| 2014 | 2,305 |
| 2015 | 2,209 |
Source from Statistics Korean, Korean Statistical Information Service (KOSIS) (http://kosis.kr).
Table 3
Prevalence, incidence, mortality of TB in some OECD countries (2012–2014) (per 100,000 population)

Democratic People's Republic of Korea (DPR Korea) is a high-risk country for TB; therefore, it is included in the 30 high-MDR-TB burden countries and 30 high-TB burden countries. It is also among ‘The three TB-high-burden country lists’ (Fig. 2, Table 3) (2). In 2015, TB incidence in DPR Korea was 561 per 100,000 individuals, which was the third highest in the world (1st: South Africa [834 per 100,000 individuals], 2nd: Lesotho [788 per 100,000 individuals]). Based on the 2016 Global Summary reported by WHO, the national BCG vaccine coverage for newborns in DPR Korea was 98% from 2010 to 2014 and 97% in 2015. These data are not completely reliable because these were not directly examined by WHO.
Figure 2
Countries with high burden of TB, MDR-TB, and TB/HIV. Countries with TB, MDR-TB, and HIV-related TB are shown as modified from the WHO Global Tuberculosis Report 2016. DPR Korea is included in both TB and MDR-TB sectors in the WHO TB high-burden country lists.
DR Congo, Democratic Republic of the Congo; UR Tanzania, United Republic of Tanzania.
DEVELOPMENT OF NEW PREVENTIVE TB VACCINES
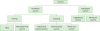
There are 2 types of TB vaccines: preventive and therapeutic (Fig. 3). The preventive vaccines include priming vaccines and boosting vaccines.
Figure 3
Classification of TB vaccines. TB vaccines were divided into preventive and therapeutic vaccines. Priming vaccines induce immune responses prior to Mycobacterium tuberculosis infection. Boosting vaccines are applied to reinforce immune responses elicited after priming. Therapeutic vaccines are used to cure TB or its symptoms with anti-mycobacterial medications.
Priming vaccines
The priming vaccine is typically administered to newborns, prior to the first exposure to M. tuberculosis. Several priming vaccines, including VPM1002 and MTBVAC, are under development.
VPM1002 (rBCGΔureC∷HLY) is a recombinant BCG mutant that expresses listeriolysin O and a hygromycin resistance marker with a deficiency of urease C (1819). Listeriolysin O is a cytolytic, pore-forming toxin protein produced by Listeria monocytogenes (2021). It increases the exposure of rBCG in the cytosol, thereby enhancing the T cell immune response through the major histocompatibility complex (MHC) class I pathway. Secreted listeriolysin O is rapidly degraded in the cytosol because of its proline, glutamic acid, serine, and threonine sequences (22). Listeriolysin O is activated at a low pH; therefore, it is designed to be deficient in urease C to guarantee an optimal pH for its activity (23). VPM1002 is being evaluated as a vaccine to replace BCG for newborns and prevent TB recurrence in adults with active pulmonary TB (24). Two phase I clinical trials (ClinicalTrials.gov identifier: NCT00749034 and NCT01113281) conducted in Germany revealed that recombinant BCG is safe for use in healthy adults of European descent (25). In addition, the stability and immunogenicity of the vaccine were confirmed (ClinicalTrials.gov identifier: NCT01113281) in 24 adults in South Africa (25). In ‘Study to Evaluate Safety and Immunogenicity of VPM1002 in Comparison With BCG in Newborn Infants in South Africa’ (ClinicalTrials.gov identifier: NCT01479972), it was discovered that VPM1002 is safe, well-tolerated, and immunogenic in comparison to the BCG vaccine for newborn infants in South Africa (25). Furthermore, a phase II clinical study titled ‘Study to Evaluate the Safety and Immunogenicity of VPM1002 in Comparison With BCG in HIV-exposed/-Unexposed Newborn Infants in South Africa’ (ClinicalTrials.gov identifier: NCT02391415) is currently underway. Phase III trials will begin in India following the phase II clinical trials in South Africa (26).
MTBVAC (ΔPhoPΔfadD26) is a live attenuated M. tuberculosis cell vaccine deficient in virulence-related genes of M. tuberculosis. The 2 deficient genes are PhoP/PhoR gene, which is involved the transcription system, and fadD26 gene, which is involved in the synthesis of phthiocerol dimycoserosate, a lipid in the bacterial cell wall. Removal of PhoP/PhoR gene leads to blockade of the ESX-1 secretion system and inhibition of the secretion of ESAT-6. Consequently, pathogenicity decreases and an immune response is induced. FadD26 is an important enzyme for glycolipid cell wall synthesis (27). The efficacy of MTBVAC is due to increased antigenicity, which is the result of increased secretion of Ag85 protein due to silencing of noncoding RNA Mcr7 (2829). There are ongoing phase II clinical trials on MTBVAC (24). Preclinical tests in rats and guinea pigs have shown that MTBVAC is a safe and immunogenic vaccine (30). A phase I dose-escalation study (ClinicalTrials.gov identifier: NCT02013245) that compared the safety and immunogenicity of MTBVAC with those of BCG vaccine was completed in February 2017. A phase Ib study (ClinicalTrials.gov identifier: NCT02729571) sponsored by Biofabri, S.L. in collaboration with the South African Tuberculosis Vaccine Initiative, Tuberculosis Vaccine Initiative, Triclinium Clinical Trial Project Management (Pty) Ltd., and Universidad de Zaragoza is yet to be conducted on MTBVAC and BCG.
Boosting vaccines
A boosting vaccine is used to enhance the immune response after a latent TB infection or a previous vaccination. It is mainly administered to adults and adolescents. Boosting vaccines may be subunit vaccines, viral-vectored vaccines, or whole-cell vaccines.
Several subunit vaccines with adjuvants such as Hybrid 1-IC31, Hybrid 1-CAF01, H56:IC31, H4 (HyVac4), M72, and ID93 (Table 4) are under development.
Table 4
Summary of the subunit vaccines with adjuvants

Hybrid 1-IC31, which contains antigen 85B (Ag85B) and ESAT-6, is used together with IC31 as an adjuvant. It induces long-lived CD4 T cell responses, resulting in the production of tumor necrosis factor (TNF)-α and IL-2 in children, regardless of M. tuberculosis infection (31). Ag85B, also known as α-antigen, is a 30-kDa mycolyl transferase protein. A hase I clinical trial (ClinicalTrials.gov identifier: NCT01049282) has been conducted to verify its safety and immunogenicity in BCG-unvaccinated volunteers with no history of TB exposure or infection. Hybrid 1-IC31 was found to be well-tolerated and safe for use in HIV-infected adults with a CD4+ lymphocyte count greater than 350 cells/mm3 in a phase II study (Pan African Clinical Trials Registry [PACTR]: PACTR201105000289276) (32). Additionally, in a phase II study recorded in the South African National Clinical Trials Register (DoH-27-0612-3947; PACTR, PACTR201403000464306), it was proven to be stable and immunogenic in individuals, irrespective of infection with M. tuberculosis. In the study, the effects of the vaccine were compared between non-infected people (QFT-negative [QuantiFERON was used to detect TB infection or latent TB infection]) and 240 QFT-positive adolescents infected with M. tuberculosis (31).
Hybrid 1-CAF01, composed of Ag85B and ESAT-6, is used together with CAF01 as an adjuvant. It increases the long-lasting T cell response (lasting 3 years), as observed by using the IFN-γ ELISpot kit. It also increases the expression of biomarkers associated with type 1 T helper (Th1) cell response, such as IFN-γ, TNF-α, IFN-γ-induced protein 10, monokine induced by IFN-γ, macrophage inflammatory protein-1β, and granulocyte-macrophage colony-stimulating factor (33).
A phase I clinical trial on Ag85B-ESAT-6 alone and a dose-escalation study on Ag85B-ESAT-6 and CAF01 in healthy adult volunteers have been completed (ClinicalTrials.gov identifier: NCT00922363). The results of the studies showed that the new liposomal adjuvant CAF01 is safe and well-tolerated (33).
H56:IC31, developed in partnership with Aeras (Rockville, MD, USA), is used for adolescents and adults. It is composed of a fusion protein comprising Ag85B, ESAT-6, and Rv2660c antigens, which are considered essential for the survival of M. tuberculosis. H56:IC31 is designed as a post-exposure vaccine that specifically targets M. tuberculosis. An open-label, dose-escalation, Phase I clinical trial (ClinicalTrials.gov identifier: NCT01967134) on AERAS-456 vaccine has been completed in 24 HIV-negative patients. This was the first-in-human clinical trial that reported no serious adverse effects, regardless of the vaccine dose administered. In addition, the vaccine was found to be safe and well-tolerated (34). Furthermore, a phase Ib clinical trial on BCG revaccination, H4:IC31, and H56:IC31 (ClinicalTrials.gov identifier: NCT02378207) has been completed in healthy HIV-1-infected adolescents.
HyVac4 (H4) was developed by the Statens Serum Institut (Copenhagen, Denmark) by using AG85B and TB10.4 as antigens, and has been combined with the adjuvant IC31 by Valneva (Lyon, France). TB10.4 is one of the ESAT-6-like proteins. In BCG-vaccinated mice, TB10.4 induces the expression of TB10.4-specific IFN-γ+ TNF-α+ IL-2+ or TNF-α+ IL-2+ CD4+ T cells (35). It is reported that, using the prime/boost regimen, HyVac4:IC31 is more immunogenic than BCG monotherapy in guinea pigs (36). Related phase I clinical trials have been completed in Europe and South Africa. Furthermore, 2 phase I dose-escalation trials (ClinicalTrials.gov identifiers: NCT02066428 and NCT02074956) on stability and immunity have been completed in BCG-vaccinated adults. Additionally, a phase II clinical trial in South Africa on the induction of polyfunctional CD4+ T cell responses and stability is underway (ClinicalTrials.gov identifier: NCT02075203).
M72 is a 72-kDa polyprotein developed by GlaxoSmithKline (Brentford, UK) and Aeras. It is a fusion protein of MTB32A and MTB39A, which are antigens of M. tuberculosis. AS01E is the adjuvant used with M72. Currently, a phase IIb efficacy study (TB-018) on M72 is scheduled to be conducted in 3,573 adult patients in 3 African countries (South Africa, Zambia, and Kenya) (ClinicalTrials.gov identifier: NCT01755598).
ID93 is a recombinant fusion protein composed of 4 MTB antigens: Rv2608, Rv3619, Rv3620, and latency-associated Rv1813 (37). Glucopyranosyl lipid adjuvant-stable emulsion (GLA-SE) acts as a Toll-like receptor (TLR) and helps to induce significant Th1 immune responses. It generates multifunctional IFN-γ+ TNF-α+ IL-2+ CD4+ T cells in both BCG-vaccinated and non-BCG-vaccinated mice and guinea pigs. A pre-clinical study that compared the effects of rifampicin or isoniazid with ID93 showed that ID93 induces pluripotent antigen-specific Th1 immune responses and inhibits M. tuberculosis-induced lung pathology in mice and cynomolgus monkeys (38). Phase IIa of the clinical trial was sponsored by the Infectious Diseases Research Institute (ClinicalTrials.gov identifier: NCT02465216). Furthermore, randomized, double-blind, and placebo-controlled clinical trials on the safety and immunogenicity of ID93 have been completed in adult TB patients who have been successfully treated with confirmed bacteriologic agents.
In addition to subunit vaccines, viral-vectored vaccines, such as MVA85A, Crucell Ad35/Aeras 402, Ad5-Ag85A, and VV-tPA-85B, have been developed as boosting vaccines.
MVA85A is a modified vaccinia Ankara (MVA) virus that expresses Ag85A. In early clinical trials on MVA85A, no significant results were observed when BCG-infected South African children (in 2013) and HIV-infected adult subjects (in 2015) were administered the vaccine as a booster (39). Moreover, a phase IIb clinical trial revealed no significant efficacy of the vaccine against M. tuberculosis infection (40) and was therefore discontinued. A phase I clinical trial (sponsored by the University of Oxford, Oxford, UK) on intradermal and aerosol formulations of the vaccine is in progress (ClinicalTrials.gov identifier: NCT02532036).
The Crucell Ad35/AERAS-402 vaccine uses a replication-defective serotype 35 adenovirus, which expresses Ag85A, Ag85B, and TB10.4. An Aeras-sponsored phase II clinical trial (Clinicaltrials.gov identifier: NCT02414828) was conducted, in which the participants were induced with a robust immune response to current or past pulmonary TB. Results of the trial revealed that treatment with the AERAS-402 vaccine is not associated with pulmonary complications (41).
The Ad5-Ag85A vaccine is formulated based on non-replicating adenovirus serotype 5 and is designed to express the M. tuberculosis antigen Ag85A. In a pre-clinical study, the vaccine was shown to improve the long-term survival of BCG-primed guinea pigs with pulmonary M. tuberculosis infection (42). Furthermore, in a McMaster University-sponsored phase I trial (ClinicalTrials.gov identifier: NCT00800670), the vaccine was found to be safe and immunogenic (43). Viral-vectored vaccines are still under development; however, recent studies have reported few drawbacks. It is reported that the prevalence of the anti-adenovirus antibody is high. Additionally, adenovirus-5-based vaccines against HIV have the potential to increase the risk of developing acquired immunodeficiency syndrome (44).
VV-tPA-85B is formulated using KVAC103, which is an attenuated vaccinia virus vector system developed at the Korea Centers for Disease Control and Prevention. It contains the Ag85B gene of M. tuberculosis H37Rv with a secretory signal peptide tPA gene. KVAC103 is the third generation of the attenuated vaccinia virus, which limits its pathogenicity. The immunogenicity and protective activity of VV-tPA-85B against M. tuberculosis has been confirmed in mice (Korea Patent Application No. 10-1749993).
Whole-cell vaccines have also been developed to boost immune responses against M. tuberculosis.
DAR-901 is a whole-cell vaccine developed at the University of Dartmouth in the United States. It is a heat-inactivated vaccine based on Mycobacterium obuense, which shares multiple antigens with M. tuberculosis for a cross-protective effect. It was developed as a booster vaccine for both HIV-infected and HIV-uninfected children and adolescents who have received BCG immunization (29). A phase I study (ClinicalTrials.gov identifier: NCT02063555) conducted in 77 HIV-negative and HIV-positive adults who had received BCG proved the safety, tolerability, and immunogenicity of the vaccine. A phase II clinical trial sponsored by Dartmouth-Hitchcock Medical Center is scheduled to be conducted on the vaccine. Participants are currently being recruited to determine if the risk of TB infection is reduced in the experimental group, which will be composed of Tanzanian adolescents who will receive BCG (ClinicalTrials.gov identifier: NCT02712424). The protective effect of the vaccine was shown to be statistically significant in a phase III clinical trial in which HIV-positive TB patients were administered multiple doses of the vaccine (45).
DEVELOPMENT OF NEW THERAPEUTIC TB VACCINES
Therapeutic TB vaccines are vaccines that are administered along with the existing drug therapy (Fig. 3) to shorten the treatment period.
RUTI, a TB vaccine candidate, is based on detoxified liposomal fragments of M. tuberculosis. It is used in addition to chemotherapy for treating latent TB infection. It was developed as an immunotherapeutic agent to reduce the amount and duration of drug therapy in patients with active TB. A phase II trial on the RUTI vaccine has been recently completed in TB patients with HIV infection (44).
Vaccae (SRL-172) is a vaccine developed by Biologic Pharmacy Co., Ltd (Longcom; Hefei City, China). It is composed of whole heat-killed Mycobacterium vaccae and has already been approved for use as a therapeutic adjuvant for patients infected with M. tuberculosis (46). A phase III clinical trial has been completed in HIV-coinfected patients. Vaccae is currently undergoing clinical trials for stability and efficacy in people with high-risk TB who are positive for the purified protein derivative skin test (ClinicalTrials.gov identifier: NCT01979900). Mycobacterium indicus pranii, formally known as Mycobacterium w, is used to prepare a whole-cell vaccine that is currently undergoing a phase III clinical trial. M. indicus pranii is a cultivable, non-pathogenic, and killed Mycobacterium species that contains antigens similar to Mycobacterium leprae (47).
ADJUVANTS FOR TB VACCINE DEVELOPMENT
Here, we briefly describe the adjuvants that have been recently used for TB vaccine development (Table 5). IC31 is an adjuvant consisting of the immunostimulatory TLR9 ligand ODN1a, an oligodeoxynucleotide, and the synthetic antimicrobial peptide H-KLKL5KLK-OH. It is used in H4 and H56 vaccines (48). AS01 contains the TLR4 ligand 3-O-desacyl-4′-monophosphoryl lipid A (MPL) and the saponin derivative QS-21 as a liposomal formulation (4950). AS01E is an adjuvant that reduces the capacity of AS01 for pediatric use. It is being used for M72 vaccine candidates. AS02 is also composed of MPL and QS-21; however, it is an o/w emulsion formulation rather than a liposomal formulation (51). CAF01 is formulated with an N,N′-dimethyl-N,N′-dioctadecylammonium (DDA) liposome and the synthetic mycobacterial immunomodulator α.α′-trehalose 6,6′-dibehenate (TDB) in a 5:1 ratio (52). It is used in H1 vaccines. GLA-SE is a stable emulsion of the synthetic TLR4 ligand glucopyranosyl lipid adjuvant, and is used in ID93 (37).

NEW APPROACHES FOR TB VACCINE DEVELOPMENT
The effect of BCG may differ depending on the route of administration. Thus, changes and improvement in the administration route must be considered in vaccine development. In animals challenged with M. tuberculosis after vaccination with BCG, the colony-forming units of M. tuberculosis in the lungs vary according to the route of administration. Aerosolized BCG administered through the nasal cavity shows the best inhibitory effect, which may be because it activates both mucosal and systemic immunity. However, it is associated with side effects, including pulmonary inflammation, which causes lung tissue damage. Oral administration of a lipid-microencapsulated vaccine formulation to mice and guinea pigs resulted in improved outcomes by inducing the production of diverse and prolonged CD4+ T cell responses. The formulation was also associated with decreased side effects. Subcutaneous injection of the vaccine stimulated delayed hypersensitivity and cytokine production by Th cells (23).
VPM1002 (rBCGΔureC∷hly) is being developed as a priming vaccine. A study of VPM1002 has been conducted in a cell model lacking a virulence gene called nuoG, which is responsible for inhibiting the apoptosis of infected host cells (53). It has been reported that BCGΔureC::hlyΔunoG shows a higher immune response and stability than BCGΔureC::hly in mice; therefore, the former may be useful as a next-generation candidate (54).
Ag85B antigen is a mycolyl transferase that is conserved in mycobacterial species such as M. tuberculosis and BCG (55). Ag85B is involved in lipid accumulation and storage, which are important processes during dormancy in M. tuberculosis. It is also used in many vaccine candidates because it induces a strong Th1 immune response. However, 8 out of the 13 vaccine candidates are subunit vaccines, and 6 of them use Ag85 antigens (Ag85A, Ag85B). Therefore, it is necessary to find novel antigens that can be used as vaccines.
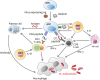
ESAT-6 is a protein encoded by a gene located in the region of difference 1, which is expressed in M. tuberculosis but is not found in BCG. It has sufficient immunogenicity in mice and humans after M. tuberculosis infection. However, inducing an immune response for the prophylactic vaccine using ESAT-6 protein requires the use of an additional adjuvant to increase the protective or vaccination effect. In a previous study, ESAT-6 showed a potent immune response when it was combined with adjuvants such as dimethyl dioctadecyl ammonium bromide and MPL (56). Furthermore, a recombinant adenovirus expressing ESAT-6 with calreticulin showed an increased immune response to ESAT-6 by secreting more IFN-γ and TNF-α; however, it could not decrease M. tuberculosis burden in the lungs (57). In addition, DNA-based vaccines prepared using the T cell epitope of M. tuberculosis are known to induce immune responses with high safety and stability (58). However, ESAT-6 itself could pose a hurdle in the process of antigen presentation. Thus, for a protective effect, modified DNA vaccines have been prepared using nano-chitosan. A previous study showed that nano-Esat-6/3eFL (nano-chitosan-based DNA vaccine prepared using fms-like tyrosine kinase 3 ligand) induced a strong immune response and exhibited a protective effect by upregulating Th1 and cytolytic T cell (CTL) responses against M. tuberculosis (59). In contrast, the Ag85A/ESAT-6 chimeric DNA vaccine induced a hypersensitivity response and worsened the severity of M. tuberculosis infection in mice (60). Recently, it was suggested that novel cell-based vaccine candidates prepared using antigen-presenting cells (APCs) such as dendritic cells and B cells can induce strong antigen-specific T cell immunity (6162). Furthermore, ESAT-6-expressing adenovirus or vaccinia virus can deliver ESAT-6 antigen to APCs, which consequently upregulate the expression of MHC and co-stimulatory molecules. However, antigen delivery using a viral vector showed dominant expression of MHC class I, and its potency of vaccine efficacy might be CD8+ T cell-dependent. Thus, an alternative method to increase MHC class II expression is required (63). Activated natural killer T (NKT) cells can increase the immunogenicity, including CD4+ T cell responses, of cell-based vaccines when α-galactosylceramide (α-GalCer) is added as a ligand (61646566). Thus, using recent approaches for the development of TB vaccines, an APC-based cell vaccine expressing ESAT-6 and loaded with α-GalCer could be formulated as a novel vaccine candidate for the prevention and treatment of M. tuberculosis infection (Fig. 4).
Figure 4
Mechanism of APC-based cell vaccines through NKT cell activation. The APC-based cell vaccine expressing antigen-triggered antigen-specific immune response. Antigens presented on APCs stimulate CD8+ T cells to express IFN-γ and sufficiently induce a CTL response against infected cells. Since antigen presentation induced by viral vectors is MHC class I-dominant, an appropriate adjuvant is required to enhance the CD4+ T cell response. α-GalCer, which is loaded on the CD1d molecule of APCs, can act as an adjuvant for immunization. NKT cells activated by α-GalCer up-regulate MHC class II molecules on APCs. Moreover, α-GalCer-loaded APCs activate NKT cells, thus allowing them to produce IL-21 to enhance the natural killer (NK) cell and CD4+ T cell response. Activated CD4+ T cells consequently accelerate antibody production in plasma cells for preventing or protecting against Mycobacterium tuberculosis. In addition to the APC-based cell vaccine, NKT cell activation using α-GalCer induces stronger immune responses by activating NKT and NK cells. As a result, the APC-based cell vaccine could be a potent candidate that functions by activating NKT cells against M. tuberculosis.
ADVANTAGES AND DISADVANTAGES OF ESAT-6 AS A VACCINE ANTIGEN
ESAT-6 forms a heterodimeric complex with CFP-10 in the phagosome lumen and is secreted through the ESX-1 secretion system (67). The heterodimer inhibits the intracellular antimicrobial response by inducing oxidative burst by downregulating the level of inducible nitric oxide synthase, which decreases the levels of nitric oxide and reactive oxygen species (68). ESAT-6 also suppresses T cell function and inhibits the intracellular defense system. In mouse neutrophils, ESAT-6 acts as a leukocidin, a bacterial enzyme that kills neutrophils and increases calcium ion influx (69). It has been suggested that a high burden of ESAT-6-expressing M. tuberculosis leads to macrophage necrosis via caspase-1- and cathepsin-B-independent pathways (70). In addition, ESAT-6 inhibits autophagy through mechanistic target of rapamycin activation, suggesting that vaccination with BCG together with ESAT-6 may increase BCG survival (7172). ESAT-6 expression is rapidly increased in host cells infected with M. tuberculosis. Furthermore, it triggers phagosomal rupture since it forms a membrane-spanning channel. ESAT-6 consists of an N-terminal, C-terminal, helix1, and helix2. The N-terminal and C-terminal are involved in cell membrane interactions, whereas helix1 and 2 are involved in membrane-spanning channel formation (73). ESAT-6 may disrupt the artificial bilayer (74) and plasma membrane of red blood cells, and play a role in the lysis of liposomes (75). In addition to causing phagosomal rupture through the formation of phagosomal membrane-spanning channels, ESAT-6 also interacts with beta 2 microglobulin (β2M), a constituent of the MHC class I molecule in all nucleated cells. ESAT-6 and ESAT-6:CFP-10 are secreted through the ESX-1 secretion system, after which they migrate to the endoplasmic reticulum in the host cell to form a complex with β2M. Consequently, cell surface expression of the MHC class I-β2M complex is inhibited and MHC-I-dependent antigen presentation is downregulated (76). M. tuberculosis infection also causes the downregulation of class II transactivator and MHC II by inducing hypermethylation of histone H3 lysine9 (H3K9me2/3). This is mediated by EsxL, one of the 23 ESAT-6 family proteins in M. tuberculosis (77).
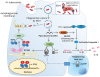
The M. tuberculosis DNA that is exposed to the cytoplasm after phagosomal rupture is recognized by cyclic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS), which induces the synthesis of cyclic GMP-AMP (cGAMP) as a cellular secondary messenger. cGAMP moves to the cytosol and activates the stimulator of interferon genes (STING) to activate TANK-binding kinase (TBK) 1 (a serine/threonine-protein kinase)-interferon regulatory factor 3 (IRF3) (Fig. 5) (7879). The activated cGAS/STING pathway finally triggers the expression of type I interferons such as IFN-α to suppress M. tuberculosis in the infected cells (8081). Additionally, TBK-1 activity and bacterial ubiquitination lead to selective autophagy of the pathogen (Fig. 5). The lipidated isoform of the microtubule-associated protein 1 light chain is involved in the ubiquitination of mycobacterial and/or phagosomal membrane fragments (82). In addition, mycobacterial DNA is recognized by absent in melanoma 2 (AIM2) and cGAS; however, NLRP3 and caspase-1 may not be involved in this process (83). It has been suggested that intracellular M. tuberculosis DNA sufficiently activates the AIM2 inflammasome, which releases IL-1β and IL-18 (84). However, another study has shown that M. tuberculosis inhibits IL-1β secretion via the ESX-1 secretion system (81). Furthermore, until date, the relationship between the cGAS and AIM2 pathways is not clear. However, it appears that components derived from M. tuberculosis, especially bacterial DNA, might play a critical role in host defense mechanisms. This consequently induces the activation of inflammasomes and the cGAS/STING pathway at the infection site to help reduce M. tuberculosis burden.
Figure 5
Mechanism of inflammatory responses in Mycobacterium tuberculosis-infected cells. In M. tuberculosis-infected host cells, the bacterium is internalized through phagocytosis. M. tuberculosis replicates and secretes its own peptides that induce pathogenic effects in the phagosome/endosome. During the life cycle of M. tuberculosis, the bacteria escape the phagosome and reach the cytosol; this process is dependent on ESAT-6, which is secreted by the ESX-1 excretion system. Cytosolic mycobacterial DNA produced by the defense mechanisms of the host cell is sensed by cGAS and AIM2. Activated cGAS uses adenosine triphosphate (ATP) and GTP to synthesize cGAMP, which acts as a secondary messenger in the host cell. cGAMP activates STING, which phosphorylates TBK-1. TBK-1 phosphorylates IRF3 and induces ubiquitination to bacteria, thus triggering autophagy. Consequently, the dimerization of phosphorylated IRF3 activates IFN-β, which suppresses the M. tuberculosis burden. On the other hand, cytosolic mycobacterial DNA is also sensed by an AIM2 DNA sensor, one of the components of the inflammasome. Activated AIM2 protein induces the protease effect of the AIM2 inflammasome, which cleaves procaspase-1 to caspase-1. Cleaved caspase-1, in turn, cleaves pro-IL-1β to IL-1β to be secreted into the extracellular space.
ESAT-6 downregulates the expression of MHC molecules and induces the migration of M. tuberculosis to the cytoplasm through host cell phagosomal rupture, which may be an adverse effect of using ESAT-6 as a vaccine. However, phagosomal rupture by ESAT-6 also induces the activation of the AIM2 inflammasome and the STING-TBK-1-IRF3 pathway, which triggers strong immune activation to eliminate M. tuberculosis infection.
CONCLUSION
In this review, we discuss the current progress in TB vaccine development. Despite the limitations of BCG, a novel TB vaccine that can replace it has not yet been developed. However, efforts to develop new TB vaccines have resulted in the production of a number of TB vaccine candidates. Here, we summarize the data on vaccine candidates that have been studied or are being investigated in clinical trials so that researchers can easily understand the current status of TB vaccine development.
We also discuss the role of ESAT-6, which is widely used as a target antigen in TB vaccine development, and question whether developing a TB vaccine with ESAT-6 is an effective strategy. ESAT-6 induces phagosome rupture in cells, which results in regulation of the migration of M. tuberculosis to the cytoplasm and is associated with the inhibition of MHC I and II. These characteristics of ESAT-6 are considered inappropriate for a TB vaccine to induce an immune response. In fact, many TB vaccine candidates that are formulated based on ESAT-6 induction have not progressed in clinical trials. In addition, problems arise when performing IGRA diagnostic tests using ESAT-6, as the results of the test cannot be attributed to the effects of the ESAT-6-based TB vaccine. To solve this problem, a study on ESAT-6-free IGRA assays is underway. In this study, EspC, EspF, and Rv2348c were screened as potent antigens and mixed with CFP-10 to produce a cocktail, which showed a diagnostic performance similar to that of QuantiFERON. As the development of vaccines using ESAT-6 proceeds, it is necessary to continue research and development in diagnosis of TB (85).
Despite the abovementioned limitations of ESAT-6, it presents several benefits as a vaccine antigen. According to previous reports on the development of ESAT-6 as a vaccine antigen, ESAT-6 is highly potent and can induce good CD4+ T cell and CTL responses to protect hosts against TB. The difficulty associated with the use of ESAT-6 appears to be the possibility of toxicity due to the immunization method. We propose the development of a novel cell-based vaccine that can overcome the low immunogenicity of ESAT-6. It appears that it is better to use a viral vector to deliver ESAT-6 antigen to APCs since ESAT-6 can suppress APC function.
Although dendritic-cell-based vaccines are well-known, we suggest the development of B-cell-based vaccines because they can sufficiently activate CD4+ T cell and CTL responses. Moreover, it is reported that an α-GalCer-loaded B cell-based vaccine induced NKT cell activation to boost immune responses elicited by a B-cell-based vaccine. Thus, a cell-based vaccine containing ESAT-6 could be a novel prophylactic and therapeutic vaccine that can enhance protection against M. tuberculosis infection.
 XML Download
XML Download