

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
ARG1
arginase 1
AZU1
azurocidin 1
C/EBP
CCAAT/enhancer binding protein
CB
cord blood
CCR3
C-C motif chemokine receptor 3
CD
cluster of differentiation
CLC
Charcot-Leyden crystal
CTSG
cathepsin G
dbcAMP
dibutyryl-cyclic adenosine monophosphate
ECP
eosinophil cationic protein
ELANE
neutrophil elastase
EoL-1
eosinophilic leukemia cell line
EPX
eosinophil peroxidase
ESGP
eosinophil-specific granule protein
GFI1
growth factor independent 1
LTF
lactotransferrin
LYZ
lysozyme
MBP1
major basic protein 1
MMP
matrix metallopeptidase
MPO
myeloperoxidase
NPGP
neutrophil primary granule protein
Olig2
oligodendrocyte transcription factor 2
PB
peripheral blood
PRTN3
proteinase 3
RNA-seq
RNA sequencing
INTRODUCTION
Neutrophils and eosinophils are 2 prominent granulocytes in the circulation that are distinguished from each other by the characteristic features of their multi-lobulated nucleus and cytoplasmic granules. The functions and roles of these granulocytes with specific features are distinctly divergent, reflecting the presence of different sets of molecular signatures. Nonetheless, they are somewhat similar in their capability to perform an array of functions including phagocytosis, destruction of microbes, mobilization of stored granules, secretion of cytokines, production of reactive oxygen species (ROS), formation of extracellular traps, regulation of other immune cell functions, and tissue damage in inflammatory conditions (12). Both types of granulocyte develop from hematopoietic stem cells via lineage-committed progenitors, each of which is driven by the combined action of a unique set of transcription factors, the cytokine milieu, and localized niches in the bone marrow (34). The eosinophil progenitor is defined by a specific combination of surface markers (5), whereas a neutrophil progenitor has not yet been described. Following commitment to lineages, the terminal differentiation pathways of the 2 granulocytes show remarkable parallelism in the temporal appearance of characteristic nuclear shape and granules through a discrete set of successive stages involving myeloblasts, promyelocytes, myelocytes, metamyelocytes, and band cells before the cells attain complete maturity.
Central to nearly all critical activities of these 2 granulocytes are their characteristic granules. Neutrophil granules are grouped into 3 types, primary (azurophilic), secondary (specific), and tertiary (gelatinase) (16), which are transcriptionally regulated and expressed during specific stages of granulopoiesis. Primary granules, including myeloperoxidase (MPO), neutrophil elastase (ELANE), proteinase 3 (PRTN3), cathepsin G (CTSG), azurocidin 1 (AZU1), and lysozyme (LYZ) are synthesized during the transition from myeloblast to promyelocyte stages. The secondary (specific) granules are dominated by a large amount of lactotransferrin (LTF), whose presence marks the beginning of the myelocyte stage, and are generated during transition from promyelocyte to myelocyte stage (67). Tertiary (gelatinase) granules are formed during myelocyte and metamyelocyte stages and serve as a repository for arginase 1 (ARG1) and matrix metallopeptidase 9 (MMP-9). Synthesis of the granule proteins during terminal differentiation is largely under the control of transcription factors, primarily CCAAT/enhancer-binding protein (C/EBP) α, C/EBPε, and growth factor independent 1 (GFI1). Expression of C/EBPα is high in myeloblasts and decreases with granulocyte differentiation, supporting its role in early granulopoiesis (8). It also directly transactivates neutrophil primary granule protein (NPGP) genes (7) and reduces proliferation by interacting with E2F (9). C/EBPε is required for transcription of granule proteins at the myelocyte stage and beyond and is indispensable for the expression of neutrophil secondary granule proteins, but not NPGPs (10). GFI1 is expressed at myeloblast and promyelocyte stages when the NPGP genes are actively transcribed. Consistent with this, GFI1 is required for transactivation of genes encoding NPGPs (1112) as well as for terminal neutrophil maturation (13).
Eosinophils have both primary and secondary granules that are acquired during development and differentiation. The primary granules store Charcot-Leyden crystal (CLC) protein (14). The eosinophil secondary granules contain 4 basic proteins called eosinophil-specific granule proteins (ESGPs): major basic protein 1 (MBP1), eosinophil peroxidase (EPX), eosinophil cationic protein (ECP), and eosinophil derived neurotoxin (EDN). However, the timing of synthesis of these ESGPs has not been described as clearly as that for neutrophil granules. Transcripts of MBP1 and EPX are already expressed in the eosinophil progenitor, together with IL-5Rα as the earliest recognizable eosinophil marker (5). Expression of these granule genes including MBP1 is driven by a combinatorial transcription factor network involving C/EBPα, C/EBPε, GATA1, and PU.1 (101516).
Using RNA sequencing (RNA-seq) analysis, we have previously demonstrated differentially expressed genes at the late stages, days 18 and 24, in an experimental model where cord blood (CB) cluster of differentiation (CD) 34+ hematopoietic stem cells were induced to differentiate toward eosinophils during a 24 day-culture period. All ESGP-encoding genes are most abundantly expressed at days 18 and 24, while numerous genes for eosinophil-specific receptors are markedly upregulated at day 24 compared with day 18 (17). A close look at the transcriptomic data revealed that transcripts for NPGPs, including PRTN3, MPO, CTSG, AZU1, and ELANE were steeply downregulated at day 24 versus day 18. While suggesting that transcripts of NPGPs might be more abundant at earlier stages of the eosinophil terminal differentiation, these results raised the question of whether eosinophils coexpress NPGPs and ESGPs in a temporal manner during terminal differentiation. Accordingly, this study aimed to examine this phenomenon by analyzing the kinetic expression of NPGPs and ESGPs and their coexpression during terminal differentiation of eosinophilic cells.
MATERIALS AND METHODS
Cell culture
The Ethics Committees of Hanyang University (HYG-11-019-1) and Soonchunhyang University Bucheon Hospital (SCHBC-2016-09-004) approved the protocols for the use of human CB and peripheral blood (PB), respectively. CD34+ cells were immunomagnetically purified from CB mononuclear cells using a MACS CD34+ MicroBead kit (Miltenyi Biotec, Auburn, CA, USA) and induced to differentiate toward eosinophils, as previously described (18). CD34+ cells were cultured in IMDM (Welgene, Gyeongsan, Korea) containing 10% fetal bovine serum (FBS), 100 U/ml penicillin, and 100 µg/ml streptomycin and supplemented with a cytokine cocktail of stem cell factor (SCF), Flt-3 ligand (Flt-3L), granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-3, and IL-5 for the first 6 days. Cells were then plated in 12-well plates in medium supplemented with IL-3 and IL-5, and cultured with 50% medium change for an additional 6 days. The cells were further cultured in medium supplemented with IL-5 for an additional 12 days, with 50% medium change every 3 days. Cell differentiation was assessed by granule formation and nucleus shape after staining with Diff-Quick (Sysmex, Kobe, Japan). PB eosinophils and neutrophils were isolated from slightly atopic individuals by centrifugation on a Percoll solution (1.070 g/ml) followed by negative selection using anti-CD16 monoclonal antibody-conjugated microbeads (Miltenyi Biotec). Their purities were greater than 95%, as evidenced by Diff-Quick staining. Eosinophilic leukemia cell line (EoL-1) eosinophilic cells were maintained in RPMI 1640 medium (Welgene) supplemented with 10% FBS, penicillin (100 U/ml), and streptomycin (100 μg/ml).
Real-time PCR
Total RNA was isolated using TRI reagent (Molecular Research Center, Cincinnati, OH, USA) and treated with DNase I (Invitrogen Life Technologies, Carlsbad, CA, USA). cDNA was reverse-transcribed from 2 μg of total RNA using SuperScript II RNase H-Reverse Transcriptase in a 20-μl reaction containing random hexadeoxynucleotide primers (Invitrogen Life Technologies), deoxynucleotide triphosphates (dNTPs) (0.5 mM), magnesium chloride (MgCl2) (2.5 mM), and DTT (5 mM). Reverse transcription was carried out at 42°C for 1 h, followed by heat inactivation at 70°C for 15 min. The resulting cDNA was amplified with the Accupower HotStart PCR Premix kit (Bioneer, Daejon, Korea). Real-time PCR was performed with SYBR Green mix (Roche, Mannheim, Germany) using a QuantStudio™ 3 Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). mRNA levels were normalized by comparison with the level of PPIA mRNA as a reference gene. Data were expressed as relative expression using the comparative cycle threshold (ΔΔCT) method. The primers used for real-time PCR are listed in Table 1.
Table 1
Real-time PCR primers

Western blot analysis
Cells were lysed in radioimmunoprecipitation assay (RIPA) buffer, resolved by SDS-polyacrylamide gel electrophoresis (PAGE), and transferred to polyvinylidene difluoride (PVDF) membranes. Blots were probed with anti-CTSG (Abcam, Cambridge, MA, USA), anti-MPO (R&D Systems, Minneapolis, MN, USA), anti-MBP1 (Abcam), anti-oligodendrocyte transcription factor 2 (Olig2) (IBL International, Hamburg, Germany), and anti-GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies, followed by incubation with anti-rabbit or anti-mouse horseradish peroxidase (HRP)-conjugated secondary Ab (Cell Signaling Technology, Beverly, MA, USA). Immunostained proteins were detected using an ECL detection system (Amersham Pharmacia Biotech Inc., Piscataway, NJ, USA).
Immunofluorescence staining
Developing CB cells were fixed in 4% paraformaldehyde, permeabilized in Tris-buffered saline (TBS) containing 0.1% saponin for 20 min, and incubated with 2% bovine serum albumin (BSA) for 1 h. The cells were stained with anti-CTSG (Abcam) and anti-human MBP1 antibody (Atlas Antibodies AB, Stockholm, Sweden) and then with Rhodamine-conjugated anti-mouse IgG antibody (Jackson ImmunoResearch, West Grove, PA, USA) or ALEXA488-conjugated anti-rabbit IgG antibody (Jackson ImmunoResearch). Isotype-matched mouse IgG2b or rabbit IgG was used as a control Ab. The cells were mounted in VECTASHIELD® Mounting Medium with DAPI (Vector Laboratories, Burlingame, CA, USA). Expression of CTSG and MBP1 was visualized using a ZEISS LSM 800 confocal microscope (ZEISS, Oberkochen, Germany) and determined using ImageJ software (National Institutes of Health, Bethesda, MD, USA) (19). For measurement fluorescence intensity, 10 cells were randomly selected at days 6, 12, 18, and 24 during differentiation of CB cells.
RESULTS
Expression of NPGP-encoding genes at late stage of differentiating CB eosinophils
We previously performed an RNA-seq analysis of differentially expressed genes at the late stages (days 18 and 24) when CB CD34+ hematopoietic stem cells were induced to differentiate toward eosinophils for 24 days (17). When we analyzed genes with reduced expression at day 24 versus day 18, the NPGP genes were enriched in the list of genes that were most downregulated (Table 2). These genes included PRTN3 (13th), MPO (14th), CTSG (27th), LYZ (35th), AZU1 (44th), and ELANE (81th), where the numbers in parenthesis indicate the rank of the most downregulated expression (the Gene Expression Omnibus database, accession number: PRJNA274725). Thus, the NPGP genes were drastically downregulated at the latest differentiation stage examined compared with the immediately preceding stage. In contrast, the transcripts of neutrophil secondary (LTF) and tertiary (MMP-2, MMP-9, and ARG1) granule protein-encoding genes remained virtually unchanged and were present at much lower abundance by 1 or 2 orders of magnitude compared with the NPGP transcripts. CD11b, which is used as an eosinophil marker (20), was upregulated as eosinophilic differentiation progressed, whereas CD16, which is not expressed by eosinophils (21), remained unchanged during these periods. Expression of GFI1 also remained unchanged.
Table 2
Expression of neutrophil-lineage genes during CB eosinophilopoiesis*

Expression of neutrophil- and eosinophil-lineage genes in developing CB cells and EoL-1 immature eosinophilic cell line
We examined the expression of NPGP genes, together with genes for ESGPs and relevant transcription factors, over the 24-day culture period by quantitative real-time PCR. Transcripts of NPGP genes, including PRTN3, MPO, CTSG, AZU1, and ELANE were considerably expressed at day 6, reached peak expression at day 12, and sharply declined at days 18 and 24 (Fig. 1). These results are consistent with the finding that expression of these NPGPs is restricted at myeloblast and promyelocyte stages (1). LYZ, which is known to be expressed in a broader window from the myeloblast to metamyelocyte stages during neutrophil terminal differentiation (1), showed extension of maximum expression to day 18 (Fig. 1). C/EBPA and GFI1 mRNA levels were highest at day 6 and decreased thereafter, thus preceding expression of NPGPs. As expected, levels of C-C motif chemokine receptor 3 (CCR3) and ECP mRNAs increased as eosinophilic differentiation progressed. MBP1 mRNA expression increased up to day 18 during CB eosinophilopoiesis and then decreased to a modestly detectable level at day 24. Low expression of MBP1 mRNA was previously demonstrated in fully mature eosinophils such as PB eosinophils (2223). C/EBPE, GATA1, and GATA2 mRNAs were progressively upregulated, as eosinophilic cells became predominant in numbers, which was commensurate with upregulation of eosinophil-specific genes. Thus, these mRNA analyses are noteworthy in that neutrophil-lineage genes display abundant expression at the early stage and then a rapid decrease from the mid stage over the course of the 24-day culture, while expression of eosinophil lineage genes progressively increases. In addition, expression of the transcription factors that regulate the lineage-specific genes kinetically precedes or parallels that of their target genes. Immature eosinophilic EoL-1 cells has been used a useful in vitro model to study human eosinophils (24). Our and other studies have previously shown that transcripts of eosinophil-specific genes including MBP1, CCR3, and IL-5 receptor alpha (IL5RA) were upregulated upon stimulation of EoL-1 cells with butyric acid and dibutyryl-cyclic adenosine monophosphate (dbcAMP) (2526). We examined whether the expression patterns of NPGP and ESGP genes were similar to those seen in differentiating CB eosinophils. Four (PRTN3, MPO, CTSG, and ELANE) of the 6 NPGP genes were abundantly present in the absence of dbcAMP but were then markedly downregulated upon the addition of this agent, while eosinophil-lineage genes (CCR3, ECP, and MBP1) were upregulated. GFI1, C/EBPA, C/EBPE, GATA1, and GATA2 transcripts remained relatively unaltered or slightly increased as EoL-1 cells differentiated (Fig. 2). Although expression of these transcription factors in the cell line was not consistent with that in differentiating CB cells, the expression patterns of genes encoding NPGPs and ESGPs were almost identical in both cultures. Therefore, it should be emphasized that the eosinophilic lineage of clonogenic nature does express NPGP genes at the immature state and then turn off these genes with concomitant increases in ESGP gene expression upon maturation. Collectively, these data clearly show coexpression of NPGP and ESGP genes in a temporal manner during terminal differentiation of the eosinophil lineage.
Figure 1
mRNA levels of neutrophil- and eosinophil-specific genes during differentiation of CB CD34+ cells toward eosinophils. Expression of 14 genes was determined by real-time PCR. The results represent the mean±standard error of the mean from 3–4 donors in which each measurement was performed in triplicate (*p<0.05 and **p<0.01, compared with day 6 CB cells).

Figure 2
mRNA levels of neutrophil- and eosinophil-specific genes in EoL-1 eosinophilic cells. EoL-1 cells were induced to differentiate in the absence or presence of dbcAMP (0.1 mM) for 1 and 3 days. Transcript levels are expressed as fold increase relative to expression at day 1 in the absence of dbcAMP. The results represent the mean±standard error of the mean of 3 to 5 independent experiments. Each measurement was performed in triplicate (*p<0.05 and **p<0.01 for comparison between the absence and presence of dbcAMP on each day).

Coexpression of CTSG and MBP1 in developing CB eosinophilic cells
We next examined whether the expression of NPGP genes was due to contamination of neutrophilic cells in the cultures of developing CB cells or whether a single eosinophilic cell changed its expression identity over the course of the 24-day culture. We performed immunofluorescence analysis over the culture period using CTSG and MBP1 as markers for neutrophils and eosinophils, respectively (Fig. 3A). At day 6, a small proportion of CB cells clearly expressed MBP1 but very weakly expressed CTSG. At days 12 and 18, the majority of CB cells robustly expressed both CTSG and MBP1, although the 2 proteins were not colocalized. At day 24 (Fig. 3A) and beyond (data not shown), MBP1 expression was maintained, but that of CTSG was markedly reduced to a relatively low level. Of note, there was also a small proportion of cells that were only CTSG-positive (see the CTSG-staining at day 24). For reference, expression of the 2 granule proteins was examined in PB eosinophils and neutrophils (Fig. 3B and C). MBP1 and CTSG were strongly detected in PB eosinophils and neutrophils, respectively, as expected. In contrast, CTSG in PB eosinophils and MBP1 in PB neutrophils were weakly detected by the immunofluorescence analysis. As control antibodies did not even produce faint signals (data not shown), these data indicate reciprocal expression of the lineage-specific protein in trace quantities that is assumed not to be expressed in eosinophils or neutrophils. These data suggest that immature eosinophils coexpress neutrophilic as well as eosinophilic granule proteins.
Figure 3
Immunofluorescence microscopy analysis of MBP1 and CTSG expression. (A) Differentiating CB cells at 4 different time points. Cells were stained with ALEXA488-anti-MBP1 (green) and rhodamine-anti-CTSG (red) antibodies. Fluorescence images were acquired using a ZEISS LSM 800 confocal microscope (ZEISS, Oberkochen, Germany), and their mean fluorescence intensities were analyzed in ten developing CB cells using ImageJ software (National Institutes of Health, Bethesda, MD, USA). These photos are representatives of developing CB cells from 3 donors (*p<0.01, compared with day 6 CB cells). (B, C) PB eosinophils (B) and neutrophils (C) were stained with the same Abs and visualized. The scale bars indicate 20 µm.

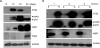
Immunoblotting analysis of CTSG, MPO, MBP1 in differentiating CB cells
For immunoblot analysis, we examined the granule proteins that had been analyzed by immunofluorescent staining. CTSG was prominently expressed at days 12 and 18, the time points displaying strong expression of CTSG by immunofluorescence. A longer exposure showed weak CTSG expression at day 6 and/or day 24 (data not shown). A similar expression pattern was observed with anti-MPO Ab (Fig. 4A), in which MPO was expressed at days 12 and 18. Of note, EPX, an ESGP, increased with a decrease in MPO. The genes MPO and EPX form a cluster on human chromosome 17 with a similar genomic structure (27), and their gene products share 70% amino acid identity (28). The anti-MPO Ab used reacted to EPX (50 kDa) as well as MPO (55 kDa), as previously demonstrated (17). In contrast, MBP1 steadily increased as differentiation proceeded, while Olig2, a late eosinophil differentiation marker (17), was detected only at day 24. PB neutrophils strongly expressed CTSG and MPO, whereas PB eosinophils expressed MBP1 and EPX, but not CTSG or MPO (Fig. 4B). Even longer exposure failed to detect CTSG and MBP1 expression in PB eosinophils and neutrophils, respectively. These results indicate that protein levels of CTSG and MBP1 in differentiating CB cells correlate with their mRNA levels and the findings from immunofluorescent staining.
Figure 4
Western blot analysis of neutrophil-specific granule proteins and ESGPs. (A) CTSG, MPO, Olig2, and MBP1 proteins were analyzed in differentiating CB cells at 6, 12, 18, and 24 days. This result is representative of cultures of CB cells from 3 donors. (B) CTSG, MPO, and MBP1 proteins were evaluated in PB eosinophils (E) and neutrophils (N) purified from 3 donors. The anti-MPO Ab used cross-reacted weakly with EPX.
DISCUSSION
In the present study, we show that CB eosinophils express NPGPs in a temporal manner during terminal differentiation. NPGPs are abundantly expressed at early and mid stages and abruptly decline as eosinophils attain full maturation, in contrast to ESGPs that show progressively increased expression. Levels of transcription factors that transactivate the NPGP- and ESGP-encoding genes kinetically correlate with expression of their target genes, being expressed prior or parallel to the respective granule protein-encoding genes. The eosinophilic expression of NPGPs is reinforced by abundant expression of NPGP transcripts in clonogenic EoL-1 immature eosinophilic cells. Upon differentiation of this eosinophilic line, the NPGP transcripts are markedly downregulated in concordance with increases in the ESGP transcripts. Our immunofluorescence analysis showed that CTSG protein, an NPGP, is expressed in almost all CB cells expressing MBP1 at intermediate stages of the terminal differentiation and is sharply downregulated in the late stage. Thus, our data strongly suggest the presence of immature eosinophils coexpressing NPGPs and ESGPs during eosinophil terminal differentiation.
Highly purified human eosinophil progenitors express MPO mRNA, albeit at a modest level, as well as transcripts of MBP1 and EPX. The eosinophil progenitor appears to be morphologically similar to cells at myeloblast and myelocyte stages, as judged by the high ratio of nucleus to cytoplasm and the round or slightly indented nucleus (5), suggesting that immature eosinophils express NPGPs. Proteomic analysis of highly purified PB eosinophils shows that the 15 most abundant proteins, including EDN, CLC, MBP1, ECP, and MBP2, account for 25% of the PB eosinophil proteome. However, many NPGPs are also present at low but non-negligible levels. Among a total of 7,086 proteins detected beyond a threshold, LYZ (72th), ELANE (976th), CTSG (1,112th), AZU (1,144th), MPO (1,181th), and PRTN3 (2,267th) are present in the PB eosinophil proteome (29). The small amounts of these NPGPs might be a vestige of abundant expression in the immature state of eosinophils. Our results are consistent with these findings, as evidenced by the finding that early differentiating CB eosinophils and the immature eosinophilic cell line substantially express nearly all NPGPs examined, which show dramatically diminished expression at more advanced stages of differentiation (Figs. 1 and 2). In addition, our data show that transcript of the transcription factor GFI1, which is considered to be specific for the neutrophil lineage (121314), is considerably expressed at the early stage (day 6) of differentiating CB cells (Fig. 1). A recent report showed that the GFI1 transcript was enriched in very early myeloid progenitors committed to eosinophils as well as to neutrophils, together with the C/EBPE transcript (30). The MPO transcript was also expressed in the early eosinophil progenitor in the same study. When eosinophils are generated from human (31) and mouse bone marrow (32) using in vitro culture systems, MPO is highly expressed at the early stage of differentiation of both cultures, although neutrophils predominate over eosinophils at that stage. Thus, these observations bolster the notion that immature eosinophils express NPGPs and relevant transcription factors. It must be stressed that, while all the studies mentioned above suggest the possibility of expression of NPGPs in immature eosinophils, this study is the first to show the coexistence of eosinophil and neutrophil granule proteins at a single cell level in differentiating eosinophils.
Interestingly, a previous proteomic analysis of PB neutrophil granules demonstrated the presence of some ESGPs. MBP1 and EPX are present virtually in primary granules of PB neutrophils, whereas CLC protein exists in secondary granules (33). As such, eosinophils and neutrophils might express NPGPs and ESGPs, respectively, in promiscuous and reciprocal manners during their terminal differentiation along a path of progenitors into fully differentiated cell types.
Functional implications of the expression of GFI1 and NPGPs in immature CB eosinophils are somewhat abstract at the present time. Of note, our results show that CB cells of an immature state (day 6 or earlier) that do not yet devote to MBP1 expression show high expression levels of GFI1 (Fig. 1). A previous study reported that forced expression of a dominant-negative form of GFI1 in neutrophilic cells or knockdown of GFI1 in eosinophilic cells results in augmented MBP1 mRNA level (34). As such, GFI1 prevents premature accumulation of MBP1, which would otherwise have negative effects on or be cytotoxic to differentiating CB eosinophils. It has been shown that some NPGPs have inhibitory effects on cytokine-driven neutrophil granulopoiesis. For instance, ELANE strongly inhibits and CTSG and PRTN3 weakly inhibit granulocyte macrophage-colony-forming units in methylcellulose culture (35). In addition, ELANE directly antagonizes granulocyte-colony stimulating factor (G-CSF)-induced proliferation of CD34+ cells by degrading G-CSF (35), which plays an essential role in proliferation, differentiation, and survival of neutrophils (36). We speculate that the production of NPGPs at early and intermediate stages might inhibit neutrophilic development during CB eosinophilic differentiation. Furthermore, NPGPs might inhibit neutrophil maturation, as supported by the finding that neutrophil secondary and tertiary granule protein transcripts are expressed at much lower levels in differentiating CB cells at days 18 and 24 (Table 2).
In summary, we found expression of NPGPs during CB eosinophilic differentiation. Almost all differentiating CB cells that express MBP1 also express CTSG. As eosinophilic differentiation progresses, expression of CTSG decreases with increased MBP1. Analysis of the transcriptomic and proteomic data from PB eosinophils and their very early progenitors suggests the possibility that immature eosinophils coexpressing both NPGPs and ESGPs exist before full maturation. Thus, the coexpression might be an inherent feature that transiently emerges during eosinophil differentiation.
 XML Download
XML Download