

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Immortality is an essential characteristic of cancer cells; our recent transcriptomic study of in vitro immortalization of human mammary epithelial cells (HMEC) reported on the epigenetic silencing of the long noncoding RNA (lncRNA) Mortal Obligate RNA Transcript (MORT; alias ZNF667-AS1) during the transition of the cells from mortal to immortal [1]. MORT is expressed in all normal, mortal human cell types analyzed to date, but is epigenetically silenced in the majority of human cancers and human cancer cell lines [1]; there is little evidence suggesting genetic mechanisms of inactivation. The experimental finding that MORT is silenced at the mortal-immortal boundary of HMEC, prior to and independent of malignant conversion, suggests that MORT silencing is an early epigenetic event in human carcinogenesis that occurs prior to malignant transformation. Since a large fraction of clinical carcinoma in situ lesions—including ductal carcinoma in situ (DCIS) [2]—show evidence of immortality in the absence of a frank malignancy [345], it is predicted that the epigenetic silencing of MORT is a common feature of this premalignant state. Breast cancer is a heterogeneous disease, but our previous analysis was not focused on MORT silencing in individual breast cancer subtypes and could not reveal if MORT silencing was an early event in clinical carcinogenesis.
Using molecular and clinical data from publicly accessible databases, we set out to evaluate the epigenetic and transcriptional status of MORT in premalignant tumors. Consistent with previous laboratory studies, our data analysis shows that DNA methylation-mediated silencing of MORT is found in both breast DCIS and colonic adenomas; this moves the timing of MORT epigenetic silencing further back to clinical premalignant conditions. The epigenetic silencing of MORT in premalignancy is maintained throughout progression towards the invasive and metastatic disease; no evidence was found for genetic mechanisms of MORT silencing. The DNA methylation-mediated silencing of MORT is significantly associated with luminal, receptor positive breast cancer; in contrast, basal triple negative breast cancers do not generally display MORT methylation or gene silencing. These early epigenetic events and the cellular mechanisms that they influence may provide new targets for cancer prevention or treatment.
DNA methylation (Illumina HumanMethylation450), gene expression (RNASeqV2), genetic (copy number variation, mutation), and available clinical data for breast tumor samples were downloaded from The Cancer Genome Atlas (TCGA). All data were analyzed with the R programming software [6] using custom scripts. MORT mean β values and mean reads per kilobase per million (RPKM) values were calculated as described previously [1]. Only nonsilent mutations were considered in the gene mutation analysis. The Spearman's rank correlation coefficient (rho) was calculated using the cor.test function. All tests between groups were performed using the wilcox.test function. When appropriate, multiple testing corrections were performed according to Benjamini and Hochberg [7]. PAM50 tumor subtype classification was used as described previously [8]. In addition to TCGA, further Illumina HumanMethylation450 datasets for breast (GSE60185 [9], GSE66313 [10], and GSE58999 [11]) and colon (GSE48684 [12] and GSE77954 [13]) cancer samples were downloaded from Gene Expression Omnibus (GEO). Where necessary, M values were converted to β values using the formula 2M/(1+2M). The reduced representation bisulfite sequencing (RRBS) and RNA-seq DCIS datasets (GSE69993 and GSE69240 [14]) were downloaded from GEO. For RRBS, the mean methylation value for the 19 covered CpGs within the MORT CpG island promoter was used for analysis.
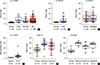
We have previously reported that MORT is epigenetically silenced by DNA methylation in breast cancer and 14 out of 16 other common human tumors investigated. To determine the potential timing of MORT silencing during human breast carcinogenesis, we analyzed the DNA methylation state of the MORT promoter in DCIS samples from two published, independent cohorts [910]. The results show a significant increase in DNA methylation of the MORT promoter in DCIS compared with normal samples (Figure 1A, B); MORT methylation did not appear to increase further during progression from DCIS to invasive breast cancer (IBC) (Figure 1A), nor during progression to the metastatic disease, as illustrated by data from a cohort [11] of pairs of primary and metastatic breast tumors (Figure 1C). A third cohort of DCIS, analyzed by RRBS and RNA-seq [14], also showed a significant increase in MORT methylation (Figure 1D) and MORT silencing in a substantial portion of the samples (Supplementary Figure 1, available online). Together, these results show that MORT silencing is an early epigenetic event that occurs during human breast carcinogenesis.
We extended our analysis of premalignant lesions and analyzed the DNA methylation state of the MORT promoter in two independent cohorts of colon adenoma samples [1213]. Although not equivalent to DCIS, colonic adenomas are early premalignant lesions in colorectal cancer and therefore might display MORT silencing if MORT silencing occurs early in carcinogenesis. The DNA methylation level of the MORT promoter in both adenoma cohorts was significantly higher than in normal colon samples; there were no further increases in MORT methylation throughout progression of the adenoma to invasive colon cancer, similar to the results for DCIS and IBC (Figure 1E, F). These results broaden the range of premalignant conditions where MORT is silenced and provide additional evidence that MORT silencing is an early epigenetic event in human carcinogenesis.
Because breast cancer is a heterogeneous disease, we sought to identify any associations between MORT loss and other clinical and molecular features of breast tumors. Figure 2A presents analysis performed to assess whether gene deletion is an additional mechanism of MORT loss in cancer, in addition to aberrant DNA methylation. The results show that MORT loss rarely occurs due to gene deletion (Figure 2A). MORT loss is therefore predominantly due to epigenetic silencing and increased DNA methylation of the MORT promoter; MORT promoter methylation is a good indicator of MORT silencing that could be used as a surrogate for MORT loss in samples where expression data are not available. When we tested the association between MORT methylation and clinical variables, we found a strong link between MORT methylation and the presence of hormone receptors. Tumors positive for individual receptors have a much higher frequency of MORT promoter methylation (Figure 2B-D) and MORT silencing (Supplementary Figure 2A-C, available online). In contrast, triple negative tumors have a low frequency of MORT methylation (Figure 2E) and near normal MORT RNA levels (Supplementary Figure 2D, available online). These data predict that MORT is predominantly silenced in luminal breast cancers; indeed, when we classified the breast tumors into subtypes according to PAM50 [15], luminal B, luminal A, and human epidermal growth factor receptor 2 (HER2)-enriched tumors had a large proportion of MORT methylation, while basal-like tumors showed no significant difference in MORT methylation or expression compared to normal control samples (Figure 2F, Supplementary Figure 2E, available online). Since some breast cancer subtypes are linked to age, we analyzed the relationship between MORT level and patient age. There is a moderate but notable increase in MORT methylation and decrease in MORT expression in tumors from patients over 60 years old (Figure 2G, Supplementary Figure 2F, available online), which is consistent with evidence of higher incidence of luminal tumors in older women [16]. Together, these results indicate that epigenetic silencing of MORT occurs predominantly in luminal, receptor-positive breast cancers.
We investigated potential associations between MORT methylation and genetic features in breast tumors. When we correlated MORT methylation with the copy number variation data on the whole genome scale, amplification of a locus containing genes CCND1 and ORAOV1 on chromosome 11 showed the strongest positive correlation (rho=0.28, adj. p=4.95e–11) with MORT promoter methylation (Figure 2H). Of these two amplified genes, only the CCND1 transcript level was significantly negatively correlated with the MORT transcript level (rho=–0.35, adj. p=8.01e–31). CCND1 is a known cancer driver linked to luminal tumors [8] and therefore its overexpression, which inactivates the RB pathway [17], might be also implicated in MORT silencing. Finally, we analyzed the links between MORT methylation and mutations in 23 genes frequently mutated in breast cancer [8]. Mutations in two of these genes were strongly linked to the level of MORT methylation. Mutation of TP53 is negatively correlated to MORT methylation (Figure 2I)—tumors carrying a TP53 mutation usually have normal levels of MORT (Supplementary Figure 2G, available online). GATA3 is frequently mutated in tumors that have reduced MORT expression (Supplementary Figure 2H, available online)—there is a positive correlation between GATA3 mutation and MORT methylation (Figure 2J). This is consistent with evidence showing that TP53 mutations are enriched in basal-like triple negative tumors, while GATA3 mutations are found predominantly in luminal tumors [8]. Overall, these findings suggest that MORT silencing in luminal breast tumors is connected to overexpression of CCND1. The lack of MORT silencing in basal tumors that frequently carry TP53 mutations indicates that MORT might act in a pathway that requires functional TP53, as the cells with nonfunctional TP53 can “afford” to express MORT and become immortal. The dissociation between TP53 mutations and MORT silencing is consistent with the in vitro immortalization model used in the discovery of MORT, where the immortal HMEC retained wild-type TP53 [18].
Overall, our analysis found that, as predicted from the in vitro model of HMEC immortalization, aberrant methylation of the MORT CpG island promoter occurs early in human breast carcinogenesis, likely at or near the point where premalignant cells gain immortality; this epigenetic silencing is maintained throughout malignant transformation and metastatic growth. In breast cancer, MORT gene silencing occurs predominantly in hormone receptor positive luminal tumors, while triple negative basal-like tumors usually show normal levels of MORT expression. Thus, taken in toto, these results are consistent with MORT silencing occurring early in breast carcino-genesis, during cellular immortalization, and before the development of luminal IBC. Delineating the molecular functions of the MORT lncRNA could provide new strategies for therapeutic or chemopreventive interventions.

 XML Download
XML Download