

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Stroke is one of the most common diseases and can induce several debilitating sequelae such as hemiplegia, aphasia, and dementia. The most important treatment for stroke thus far is thrombolysis, ideally within the so-called golden hour during which treatment is most likely to be effective, or at least within around 4.5 hours of symptom onset of a cerebral infarction.1 However, not all patients with a cerebral infarction are candidates for thrombolysis. Due to the risk of hemorrhage, there are strict inclusion and exclusion criteria for thrombolytic therapy, and because of the nature of the disease and its treatment, many patients must unfortunately live with long-term neurologic deficits. Numerous clinical trials have tested diverse putative neuroprotective agents and stem cells as treatments for these deficits, but almost all of them have been found to be ineffective. Therefore, continued effort is necessary to develop new therapeutic strategies for treating cerebral infarctions.
Ways of reversing the sequelae of cerebral infarction have been investigated, and various molecular pathways have been found to play critical roles in the pathogenesis. Based on these findings, several methods for blocking the pathogenic mechanisms have been proposed and developed. One pathogenic mechanism is the pathway involving phosphatidylinositide 3-kinase (PI3K), which has been investigated intensively.234 Since this pathway is important in cell survival and is significantly affected by ischemia, many attempts have been made to modulate it for the treatment of cerebral infarction and to prevent the programmed cell death caused by ischemic strokes.
Endogenous neural stem cells (NSCs) are well known to exist in the subventricular zone of the ventricles and subgranular zone of the hippocampus.56 They can proliferate under appropriate stress and then differentiate into various neuronal cells. Several studies have found that mild-to-moderate ischemic strokes can induce endogenous neurogenesis, and this neurogenesis may function to reduce the severity of neurologic sequelae.789 Therefore, methods for increasing endogenous neurogenesis have been sought in numerous studies. It has been established that the role of the PI3K pathway is crucial in the function of NSCs; therefore, if the PI3K pathway can be activated in NSCs after cerebral infarction, it could contribute to the recovery of subsequent neurologic deficits.
This review describes the PI3K pathway itself, its roles in the brain and in the activity of NSCs after cerebral infarction, and the possibility of using methods for activating the PI3K pathway in NSCs for the treatment of cerebral infarction.
WHAT IS THE PI3K PATHWAY?
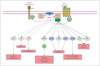
It is well known that the PI3Ks are involved in various cellular functions, such as cell proliferation, growth, differentiation, motility, survival, and intracellular trafficking. The PI3K pathway is necessary for the survival of both neurons and NSCs.1011 PI3K is not a single enzyme, but rather a family of many different subtypes. The PI3K family is divided into three different classes (Class I, Class II, and Class III) based on the primary structure, regulation, and in vitro lipid substrate specificity.12 Among these, the Class I PI3Ks are the best understood and can be divided into two groups: Class IA (p110α, p110β, and p110δ) and Class IB (p110γ).13 The PI3K pathway is known to interact with the insulin receptor substrate (IRS) and is closely linked with the tumor suppressor phosphatase and tensin homolog (PTEN), which inhibits PI3Ks. When the IRS is activated by insulin, it activates PI3Ks and then regulates glucose uptake through diverse phosphorylation events. In detail, activated PI3Ks phosphorylate the 3-position hydroxyl group of the inositol ring of phosphatidylinositol (Ptdlns) to produce phosphoinositide Ptdlns(3,4,5)P3 from Ptdlns(4,5)P2.1415 Ptdlns(3,4,5)P3 activates many different downstream effectors, the most well known of which is Akt (protein kinase B).16 Akt phosphorylated by PI3Ks (pAkt) affects many important downstream signals, including mouse double minute 2 homolog (MDM2), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), endothelial nitric oxide synthase (eNOS), mammalian target of rapamycin (mTOR), and S6 kinase, and inhibits Forkhead box O (FOXO)s, BAD, and glycogen synthase kinase (GSK)-3β.1617 These effects contribute to growth, translation, and cell-cycle regulation, glucose metabolism, DNA repair, and inhibition of apoptosis (Fig. 1).1617
Many chemicals affecting the PI3K pathway have been developed and investigated for the possibility of clinical applications. In particular, PI3K inhibitors are being investigated for the treatment of various cancers such as breast cancer and colorectal cancer,18 and PI3K activators including an IRS-1 substrate and platelet-derived growth factor receptor 740Y-P (PDGFR740Y-P) have been produced and used to enhance neuronal cell survival and differentiation.192021
ROLE OF THE PI3K PATHWAY IN THE BRAIN AFTER CEREBRAL INFARCTION
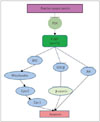
The protective role of the PI3K pathway in the brain after cerebral infarction has been emphasized. It has been reported that ischemia and reperfusion both inhibit the PI3K pathway and induce apoptosis.23 On the other hand, activation of the PI3K pathway after ischemic injury has also been reported.422 Considering our previous findings19 and those of other investigators,23422 this discrepancy may be attributable to (among other possibilities) differences in evaluation time after ischemia or reperfusion injury, ischemic duration, or the use of different cell types such as neurons or astrocytes. The level of pAkt has been found to decrease in the hyperacute phase (less than 2 or 3 hours) after ischemic events, is slightly increased in the acute phase (from 2 or 3 hours to several hours), and then decreases 6-12 hours after an ischemic event.1923 In summary, ischemia or reperfusion inhibits the PI3K pathway and then induces cell death after ischemic injury. Ischemia or reperfusion inhibits PI3K and then reduces the phosphorylation of Akt. Decreased pAkt cannot inhibit BAD, caspase-3, or GSK-3β, for example, and these changes are associated with cell death after ischemia (Fig. 2).
These findings have prompted many studies demonstrating that activation of the PI3K pathway can contribute to the protection of neurons and the brain from ischemic injury. For example, vascular endothelial growth factor protects the brain after focal cerebral ischemia through activation of the PI3K pathway;24 the Ang1-Tie2-PI3K (angiopoietin-1-tunica intima endothelial kinase-2-PI3K) axis initiates survival responses in neural progenitor cells after oxygen and glucose deprivation;25 and humanin enhances the PI3K pathway and then contributes to protecting the brain against cerebral ischischemia and reperfusion injury.26 Together these findings indicate that appropriate activation of the PI3K pathway may be useful for promoting brain cell survival and thus reducing cell death after stroke.
ROLE OF THE PI3K PATHWAY IN NSCS
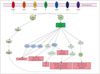
NSCs are very important in the regeneration of brain tissue that has been damaged by cerebral infarction. It has been established that endogenous NSCs are located in the subventricular zone of the lateral ventricles and the subgranular zone of the hippocampus, and that they contribute to neurogenesis.56 Endogenous NSCs can differentiate into neurons, astrocytes, and oligodendrocytes in the presence of exogenous stimuli from their environment.27 It has also been reported that the role of the PI3K pathway is very important for the activity of NSCs. The PI3K/Akt pathway controls the proliferation, differentiation, and migration of endogenous NSCs.28 For example, several kinds of neurotrophic factors such as brain-derived neurotrophic factor (BDNF),29 fibroblast growth factor (FGF),28 transforming growth factor β (TGF-β),30 insulin-like growth factor-1 (IGF1),28 and C-X-C motif chemokine 12; also known as stromal-cell-derived factor 1α (SDF-1α) (CXCL12)31 activate the PI3K pathway, and activated PI3K/Akt affects mTORC1,32 Ras-related C3 botulinum toxin substrate 1 (Rac1),33 and Rho-kinase (ROCK)34 in NSCs.
In more detail, SDF-1α binds to C-X-C chemokine receptor type 4 (CXCR4) and activates PI3K, which in turn increases the phosphorylation of Akt-1.31 pAkt-1 induces the phosphorylation of the transcription factor FOXO3a,31 which contributes to the proliferation of NSCs.31 It has also been reported that activation of the PI3K and extracellular-signal-regulated kinase (ERK) pathways markedly enhances the brain-ischemia-induced proliferation of NSCs.35 A link between Akt and cAMP response-element-binding protein (CREB) also plays important roles in the proliferation of NSCs.28
The role of Akt in NSC differentiation remains a matter of controversy. Wang et al.36 and Zhang et al.37 reported that Akt mediates proneuronal basic-loop-helix transcription factor neurogenin 1 and another transcription factor, Brn-4, and that these factors regulate neuronal differentiation and neurite outgrowth. However, Jin et al.38 and Chan et al.39 suggested that the PI3K pathway is not required for the differentiation of NSCs. Therefore, further study is required to define the exact role of the PI3K pathway in the differentiation of NSCs.
Moreover, migration is one of the most important characteristics of NSCs. Activation of the receptor tyrosine kinase ErbB4 leads to activation of PI3K, and in turn increases the migration of NSCs.40 Li et al.41 showed that up-regulation of CXCR4 activates the PI3K pathways and then enhances the migration of NSCs toward SDF-1α.
Based on all of the above findings, it can be concluded that the PI3K pathway plays important roles in the proliferation, differentiation, and migration of NSCs (Fig. 3). In addition, there is indirect evidence supporting the importance of the role of the PI3K pathway in NSC activity. Groszer et al.42 reported that PTEN deletion directly induces activation of the PI3K pathway, thus increasing the proliferation of NSCs and decreasing their death in the subventricular zone. Thus, ultimately, the PI3K pathway plays key roles in the proliferation, differentiation, migration, and survival of NSCs.
ROLE OF THE PI3K PATHWAY IN NSC ACTIVITY AFTER CEREBRAL INFARCTION
It is clear that ischemia induces neuronal cell death in the brain via diverse mechanisms.43444546 However, it is interesting to note that a small amount of ischemia, such as that resulting from a transient cerebral ischemic attack, is also capable of inducing neurogenesis.4748 Ischemia transiently increases BDNF and nerve growth factor (NGF) in the brain.49 Based on the findings that epidermal growth factor (EGF), FGF-2, and BDNF augment the proliferation of NSCs and their differentiation to mature neurons in the brain,505152 it was thought that ischemia could transiently increase the expression of neurotrophic factors in the brain and then induce neurogenesis via the activation of endogenous NSCs. This hypothesis was indirectly supported by the finding that intraventricular injection of FGF-2 or EGF after brain ischemia increased the number of NSCs in the hippocampus.5354 However, it has not yet been established how these factors enhance neurogenesis in brain tissue that has been damaged by ischemia.
As described in the sections above, those neurotrophic factors secreted in response to ischemia strongly stimulate receptor tyrosine kinases and then activate PI3K/Akt. Activated Akt plays several crucial roles in the stimulation of adult neurogenesis from NSCs. This is supported by the findings that PI3K inhibitors such as LY294002, wortmannin, and PD98059 block DNA synthesis in NSCs and inhibit their proliferation.1155
In addition, the migration of NSCs to the lesion after ischemia is important for the regeneration of the damaged brain tissue. The PI3K pathway plays a critical role during the process of migration. PI3K/Akt activated by erythropoietin enhances the secretion of matrix metalloproteinase (MMP)-2 and MMP-9, which are very important in the migration of NSCs.56 This was reconfirmed by the finding that MMP inhibitors significantly reduce the migration of neuroblasts in the brain damaged by transient focal cerebral ischemia.57
A small degree of ischemia, such as that resulting from a transient cerebral ischemia, may potentiate NSC proliferation, differentiation, and migration. However, severe ischemia induces endogenous NSC damage, and a recent study showed that the clinical improvement after treatment with mesenchymal stem cells (MSCs) was associated with the degree of involvement of the subventricular zone of the lateral ventricle, which is well known to have a relatively high concentration of NSCs. This suggests that the treatment effect of MSCs is accentuated due to a smaller contribution to the repair by damaged endogenous NSCs.58 Given that prolonged and severe hypoxia inhibits the PI3K pathway, it is possible that large and severe infarcts inhibit the pathway in NSCs and then induce the NSC death.
PI3K PATHWAY ACTIVATION TO ENHANCE RECOVERY OR REGENERATION OF DAMAGED BRAIN TISSUE AFTER A CEREBRAL INFARCTION
The findings described so far suggest that the PI3K pathway plays several crucial roles in neurogenesis by activating the proliferation, migration, and differentiation of NSCs. Recent studies have shown that the PI3K pathway mediates neurogenesis of NSCs via various downstream activators.596061 In detail, PI3K activates mTORC2 in NSCs and then enhances their survival, proliferation, and neurogenesis.59 Inactivation of GSK-3 directly by PI3K induces mothers against decapentaplegic homolog 1 of Sma (SMAD-1) and then contributes to axonal regeneration after axotomy.6061 In addition, several chemicals, drugs, and neurotrophic factors have been reported to activate PI3K (Table 1).62 636465666768
It is possible that appropriate activation of the PI3K pathway and enhanced endogenous neurogenesis with these activators may help the regeneration of brain tissue damaged by an ischemic stroke and the recovery of neurobehavioral functions impaired by cerebral infarction. There is a significant body of evidence indirectly supporting this hypothesis. For example, repetitive ischemic preconditioning inhibits brain damage after focal cerebral ischemia via activation of the PI3K pathway,69 heme oxygenase-1 (HO-1) protects hippocampal neurons from ischemic stroke via activation of the PI3K pathway,70 and formononetin activates the PI3K pathway and prevents ischemia or reperfusion injury of the brain.71
These findings suggest that direct activation of PI3K can contribute to protecting the brain after cerebral infarction and that NSCs play an important role in this protection, although there is no direct supporting evidence for this yet. From the therapeutic perspective, early activation of the PI3K pathway might be useful for protecting the brain in the hyperacute and acute stages of ischemic stroke, and its delayed activation would be helpful for restoring the ischemia-damaged brain in the subacute and early chronic stages. Further studies demonstrating the relationship between direct activation of the PI3K pathway, the protection of the brain after cerebral infarction, and regeneration by NSCs are necessary to confirm these hypotheses.
CONCLUSIONS
The PI3K signaling pathway plays several crucial roles in the survival, proliferation, differentiation, and migration of NSCs. This pathway also contributes to the protecting the brain after cerebral infarction following stroke. Further investigation into methods for enhancing endogenous neurogenesis via activation of the PI3K pathway is warranted.

 XML Download
XML Download