

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Most thyroid diseases, including autoimmune thyroiditis and thyroid cancer, have been recognized to have high heritability [12]. In twin studies, a high concordance rate for Graves' disease (GD) in monozygotic twins was reported, in the range of 50% to 70%, compared with 3% to 25% in dizygotic twins [13]. A study of autoimmune hypothyroidism likewise showed a 55% concordance in monozygotic twins [4]. Familial clustering of autoimmune thyroid disease has been consistently reported [567]. Hemminki et al. [7] showed that the familial standardized incidence ratios for GD were 4.49 for individuals with an affected parent, 5.04 for those whose singleton sibling was affected, 310 when two or more siblings were affected, and 16.45 in twins. For Hashimoto's thyroiditis (HT), the sibling risk ratio was 28 based on data from the National Health and Nutrition Examination Survey III [8], and a similar risk was confirmed in data from Germany [5]. These pieces of evidence suggest the existence of a genetic predisposition to autoimmune thyroid diseases.
Thyroid function, including levels of thyroid hormone and thyroid stimulating hormone (TSH), is regulated within a narrow range in individuals, although the inter-individual variability is large [9]. This suggests that every individual has his or her own set point of thyroid function [10]. About 40% to 60% of variation in thyroid function has been estimated to be determined by genetic factors [101112]. Thyroid cancers also show a high degree of heritability, with genetic factors accounting for more than 50% of the causes of thyroid cancer [2]. Except for medullary thyroid cancer, which is well known to be caused by germline or somatic mutations, the prevalence of familial differentiated thyroid cancer (DTC) accounted for 2.5% to 11.3% cases of DTC [1314151617]. Only 5% of cases of nonmedullary familial DTC were reported to be of the syndromic form, which is accompanied by well-known germline mutations, including Cowden syndrome, familial adenomatous polyposis, Gardner syndrome, Carney complex type 1, Werner syndrome, and DICER1 syndrome [18]. Thus, the majority of cases of familial DTC were found not to be caused by germline mutations, despite its pattern of genetic inheritance.
Thus, genetics plays a prominent role in most thyroid-related phenotypes. Research into the genes responsible for thyroid disease has identified several candidates [19]. However, candidate gene studies have been controversial and have shown very few reproducible findings. Panicker [19] published a thorough review of genetic studies of thyroid function and autoimmune thyroid diseases conducted through 2010. In the last decade, genome-wide association studies (GWAS) have been extensively used to identify genes involved in complex diseases [20]. GWAS have facilitated the screening of a large proportion of the genome and discovered a variety of susceptibility genes. GWAS have been widely applied in autoimmune thyroid diseases, thyroid function, and thyroid cancer, and have identified susceptibility genes for thyroid-related phenotypes. Herein, we comprehensively review the wide range of discoveries from GWAS conducted in Western and Asian populations regarding autoimmune diseases, thyroid function, and thyroid cancer.
GWAS FOR AUTOIMMUNE THYROID DISEASES
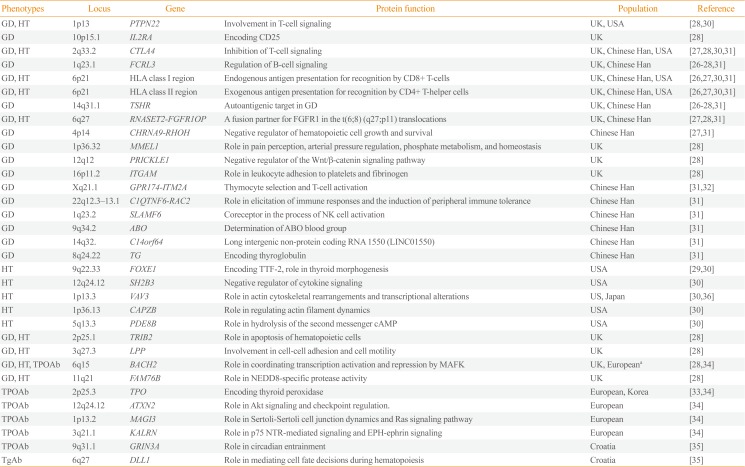
Several candidate gene studies identified putative susceptibility variants for GD, but only the human leukocyte antigen (HLA) locus and the cytotoxic T-lymphocyte associated protein 4 (CTLA4), thyroid stimulating hormone receptor (TSHR), and protein tyrosine phosphatase, non-receptor type 22 (PTPN22) loci were confirmed in subsequent replication studies [2122232425]. The first genome-wide analysis using 14,436 nonsynonymous single-nucleotide polymorphisms (SNPs) for GD was performed by the Wellcome Trust Case Control Consortium, and showed that three loci (HLA, TSHR, and Fc receptor like 3 [FCRL3]) were associated with GD [26]. A subsequent GWAS with >500,000 SNPs confirmed previously reported loci and identified a novel region of susceptibility loci at 6q27 (the ribonuclease T2 [RNASET2]-FGFR1 oncogene partner [FGFR1OP]-CCR6) and an intergenic region at 4p14 (GDCG4p14) [27]. Several GWAS of autoimmune thyroid diseases (GD, HT, and positivity of anti-thyroid peroxidase [TPO] antibody or anti-thyroglobulin [Tg] antibody) and hypothyroidism have further identified susceptibility loci (Table 1) [2627282930313233343536]. Since GWAS of HT have been performed for a variety of phenotypes including self-reported hypothyroidism, biochemical hypothyroidism with positive antibodies, antibody positivity, and level of antibodies, caution is needed when interpreting the results. Several types of hypothyroidism might not have an autoimmune etiology, and autoimmunity does not necessarily lead to hypothyroidism. Thus, careful consideration regarding the phenotype is required when interpreting the biological mechanisms of the associated genes identified through GWAS of autoimmune thyroid diseases.
A heterogeneity analysis between GD and HT showed that GD and HT share several susceptibility loci (HLA, PTPN22, and CTLA4), while an association with TSHR was exclusively seen in GD patients. The majority of genes associated with autoimmune thyroid disease are thought to play a major role in autoimmune processes, including disrupted T-cell regulation and peripheral immune tolerance [37]. Variants in thyroid-specific loci, including TSHR and forkhead box E1 (FOXE1), could affect the immune recognition of autoantigens and antibody generation [37].
GWAS OF THYROID FUNCTION
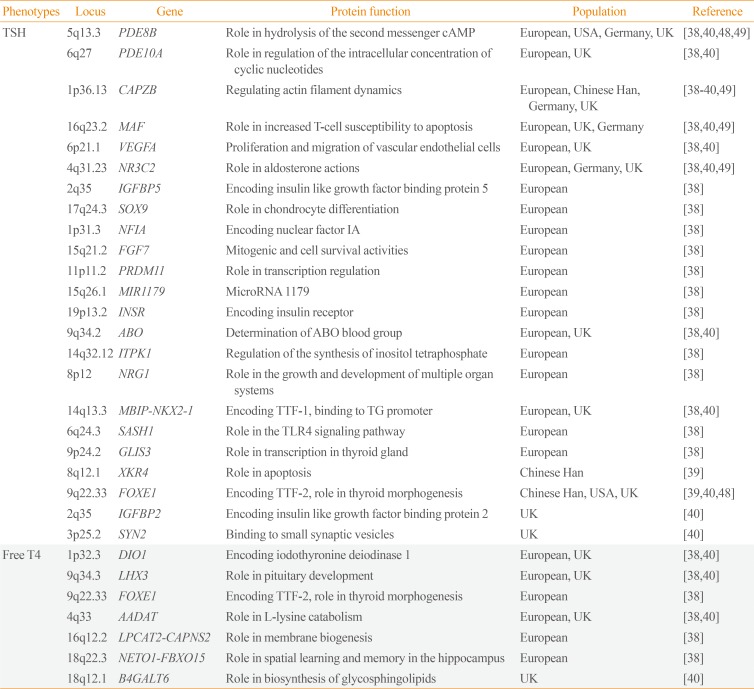
Thyroid function, including levels of free thyroxine (T4) and TSH, is highly heritable even in euthyroid subjects. A large meta-analysis of GWAS of serum levels of TSH and free T4, in 26,420 and 17,520 euthyroid European individuals, respectively, was performed, identifying many susceptibility loci for levels of TSH (phosphodiesterase 8B [PDE8B], phosphodiesterase 10A [PDE10A], capping actin protein of muscle Z-line subunit beta [CAPZB], MAP, vascular endothelial growth factor A [VEGFA], nuclear receptor subfamily 3 group C member 2 [NR3C2], insulin like growth factor binding protein 5 [IGFBP5], SRY-box 9 [SOX9], nuclear factor I A [NFIA], fibroblast growth factor 7 [FGF7], PR/SET domain 11 [PRDM11], microRNA 1179 [MIR1179], insulin receptor [INSR], ABO, inositol-tetrakisphosphate 1-kinase [ITPK1], neuregulin 1 [NRG1], MAP3K12 binding inhibitory protein 1 [MBIP], SAM and SH3 domain containing 1 [SASH1], and GLIS family zinc finger 3 [GLIS3]) and levels of free T4 (iodothyronine deiodinase 1 [DIO1], LIM homeobox 3 [LHX3], FOXE1, aminoadipate aminotransferase [AADAT], lysophosphatidylcholine acyltransferase 2 [LPCAT2]/calpain small subunit 2 [CAPNS2], neuropilin and tolloid like 1 [NETO1]/F-box protein 15 [FBXO15]) [38]. A GWAS of TSH levels was also conducted in 1,346 Chinese Han individuals [39]. Zhan et al. [39] confirmed previously reported TSH susceptibility loci near FOXE1 and CAPZB and identified novel variants in XK related 4 (XKR4). Whole-genome sequence-based analysis was performed to examine the genetic architecture for levels of free T4 and TSH, and further identified novel variants on synapsin II (SYN2), PDE8B, and beta-1,4-galactosyltransferase 6 (B4GALT6) [40]. They also found a rare functional variant (minor allele frequency=0.4%) in the transthyretin (TTR) gene, which is located near B4GALT6. This study showed that common variants explained over 20% of the variance in TSH and free T4 and that a substantial amount of heritability of thyroid function could be explained by rare variants with larger effects. Results of GWAS for thyroid function are summarized in Table 2.
Thyroid function may be affected by the presence of antibodies to TPO or Tg, even in the normal range. In GWAS of thyroid function, data on the presence of antibodies were limited. Therefore, it is difficult to conclude that the genes found in GWAS of thyroid function determine an individual set point of the hypothalamus-pituitary-thyroid axis. Several genetic loci identified in GWAS of thyroid function were also found in GWAS of autoimmune thyroid diseases (FOXE1, CAPZB, and PDE8B). A detailed examination of the presence of antibodies should be considered when performing GWAS of thyroid function in the future. In addition, only very limited GWAS of thyroid function have been performed in Asians, so more research is needed.
GWAS OF THYROID CANCER
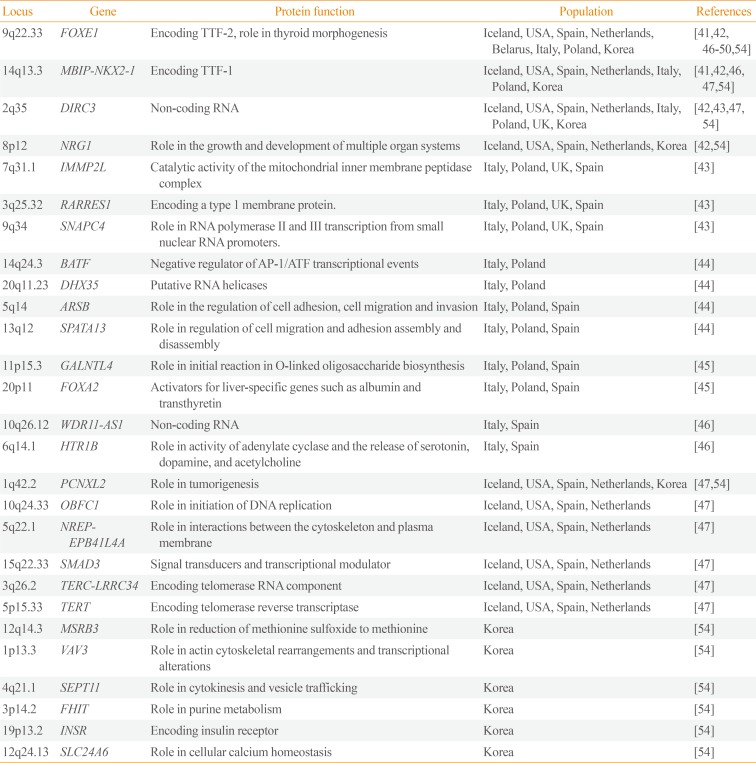
The first GWAS of thyroid cancer was reported in 2009 and showed that common variants located on 9q22.33 (FOXE1) and 14q13.3 (NK2 homeobox 1 [NKX2-1]) were associated with DTC [41]. Associations at FOXE1, MBIP/NKX2-1, disrupted in renal carcinoma 3 (DIRC3), and NRG1 have been identified and repeatedly confirmed in individuals of European ancestry [41424344]. Several markers associated with DTC, including inner mitochondrial membrane peptidase subunit 2 (IMMP2L), retinoic acid receptor responder 1 (RARRES1), small nuclear RNA activating complex polypeptide 4 (SNAPC4), basic leucine zipper ATF-like transcription factor (BATF), DEAH-box helicase 35 (DHX35), UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase-like 4 (GALNTL4), 5-hydroxytryptamine receptor 1B (HTR1B), forkhead box A2 (FOXA2), and WDR11 antisense RNA 1 (WDR11-AS1), were identified but not replicated in other studies [43444546]. A recent meta-analysis of GWAS including a total of 3,001 DTC patients and 287,550 controls from five study groups of European populations found five novel loci (pecanex-like 2 [PCNXL2], telomerase RNA component [TERC], neuronal regeneration related protein [NREP]-erythrocyte membrane protein band 4.1 like 4A [EPB41L4A], oligosaccharide-binding folds containing 1 [OBFC1], and SMAD family member 3 [SMAD3]) [47]. Table 3 provides the susceptibility loci identified in GWAS of thyroid cancer [3839404849]. The most robust signals were detected on 9q22.33 (FOXE1) in Caucasians [4150]. The FOXE1 locus was also reported to be a susceptibility gene for radiation-related thyroid cancer [50]. A functional study showed that common variants on FOXE1 regulated FOXE1 transcription through the recruitment of the upstream stimulatory factor 1 (USF1)/USF2 transcription factors [51]. Several reports demonstrated that variants of FOXE1 were related to aspects of the clinical aggressiveness of papillary thyroid cancer (PTC), such as tumor stage, size, lymphocytic infiltration, and extrathyroidal extension [5253].
Recently, we reported 15 variants from 11 loci associated with DTC in a Korean GWAS including 1,085 cases of DTC and 8,884 controls [54]. The most robust signals were detected in the NRG1 gene, and expression quantitative trait loci analysis showed that variants on NRG1 were also associated with NRG1 expression in thyroid tissues [54]. He et al. [55] also showed that the expression levels of NRG1 isoforms were significantly correlated with genotypes. NRG1 encodes neuregulin-1, which acts on the erb-b2 receptor tyrosine kinase (ERBB) family of tyrosine kinase receptors. In a study of the intrinsic resistance of PTC to a B-Raf inhibitor, ERBB2/ERBB3 activation was found to be dependent on autocrine production of neuregulin-1 [56]. NRG1 dysregulation is also closely related with the phosphoinositide 3-kinase (PI3K)-AKT and mitogen-activated protein kinase (MAPK) signaling pathway via ERBB [57]. Our gene set enrichment analysis data showed that variants on NRG1 were associated with many pathways related to cellular growth or cancer, and the ERBB-MAPK signaling pathway was the most significantly enriched. This evidence indicates that NRG1 expression in thyroid tissue could contribute to increased DTC risk via ERBB signaling.
Our results confirmed previously reported loci (FOXE1, NKX2-1, DIRC3, and PCNXL2) from GWAS of European populations and found novel susceptibility loci (vav guanine nucleotide exchange factor 3 [VAV3], INSR, MRSB3, fragile histidine triad [FHIT], septin 11 [SEPT11], and solute carrier family 24 member A6 [SLC24A6]) associated with DTC. Specially, a variant of SLC24A6 was associated with a specific risk of follicular thyroid cancer, for which the genetic factors that increase the risk of thyroid cancer may vary depending on the cancer subtype. Signals on VAV3, INSR, MRSB3, FHIT, SEPT11, and SLC24A6 were only identified in Koreans, suggesting between-study heterogeneity in GWAS of DTC.
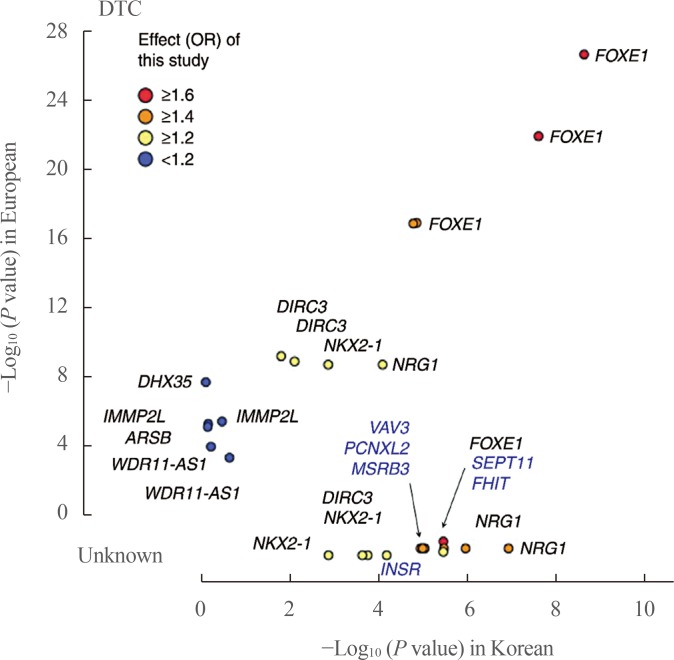
In GWAS in European and Korean populations, some genetic loci (FOXE1, NKX2-1, DIRC3, NRG1, and PCNXL2) were commonly found, while certain susceptibility loci were only found in either the European or Korean population. In addition, the risk allele frequency of commonly found SNPs differs by race, and the DTC risk by genotype varies across ethnicities. For example, the risk allele frequencies of variants on FOXE1 were reported to be 0.14 to 0.34 in Europeans and 0.08 to 0.13 in Asians, suggesting ethnic differences in allele frequencies and a small genetic contribution of variants on FOXE1 to the development of DTC in East Asians [58]. Moreover, common variants on FOXE1 were associated with an increased risk of DTC, with an odds ratio (OR) of 1.80 in the European population, but the OR was 1.35 in East Asians [58]. A comparison of these associations, including effect size (OR) and P values, between Europeans and Koreans is shown in Fig. 1 [54].
CONCLUSIONS
Twin and family studies of autoimmune thyroid diseases and thyroid cancer have indicated high heritability, suggesting that genetic factors play a key role in disease onset. Previous candidate-gene studies have limitations, such as lack of reproducibility and small sample sizes with limited statistical power. In the last decade, GWAS have unraveled the many forms of genetic predisposition to autoimmune thyroid disease, thyroid function, and thyroid cancer. These genetic discoveries provide insight into the pathogenesis of these diseases and provide opportunities to develop new therapies.
 XML Download
XML Download