

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Glucagon-like peptide 1 (GLP-1) is a 30-amino-acid incretin hormone produced by intestinal L-cells. Although it is well known to exert beneficial effects by lowering postprandial glucose levels, GLP-1 has been reported to have multiple other functions, including modulation of cell proliferation, differentiation, and apoptosis in various tissues [1]. Many recent studies reported an anabolic effect of GLP-1 on bone, with one study showing that GLP-1 reversed hyperlipidemia-related osteopenia in a rat model [2]. Another study reported an insulin-independent anabolic effect of GLP-1 in an insulin-resistant rat model [3]. However, the exact mechanism of this effect has not been established. Results concerning expression of the GLP-1 receptor (GLP-1R) in osteoblastic cells are inconsistent. GLP-1R expression has been reported in various osteoblastic cell lines, albeit at differing levels [4,5]. Adipose-derived stem cells (ADSCs) are reported to be multipotent and capable of osteogenic differentiation [6,7,8], and studies suggest that they could be an abundant, accessible, and replenishable cell source for bone cell therapy applications [9,10]. In an animal study, critical-size mouse calvarial defects were healed using scaffolds seeded with ADSCs [11].
To our knowledge, no studies have described the expression of GLP-1R in ADSCs. The aim of this study was to evaluate the expression of GLP-1R during the osteogenic differentiation of ADSCs.
METHODS
Design and participants
ADSCs were isolated from subcutaneous abdominal adipose tissue obtained from three male donors (mean age, 40 years) during plastic surgery. Body mass indices of the donors were 22.6, 26, and 23.6 kg/m2. They had received no medications, including antilipidemic or antidiabetic agents. The donors provided written informed consent, and the study protocol was approved by the Institutional Review Board of Pusan National University Hospital (IRB number 2013-8).
Isolation and culture of cells
Knife biopsies of adipose tissue were immediately placed in minimum essential medium-alpha (MEM α, Life Technologies, Carlsbad, CA, USA) supplemented with 100 U/mL penicillin and 100 µg/mL streptomycin (Life Technologies). Samples were transported to the laboratory and processed within 30 minutes of excision. Using a sterile technique, the tissue was finely minced and digested with 0.075% type I collagenase (Sigma-Aldrich, St. Louis, MO, USA) for 30 minutes at 37℃, with vigorous shaking. Then, 25 mL of MEM α containing 10% fetal bovine serum (FBS) were added to neutralize the collagenase, and the suspension was centrifuged at 3,000 rpm for 10 minutes. Next, samples were filtered through a 70µm nylon cell strainer (BD Biosciences, San Diego, CA, USA), washed with phosphate-buffered saline (PBS), and centrifuged at 1,600 rpm for 10 minutes. The isolated ADSCs were maintained and expanded in ADSC culture medium consisting of MEM α containing 10% FBS, 100 U/mL penicillin, and 100 µg/mL streptomycin at 37℃ in 5% CO2. This initial culture was referred to as passage zero. The medium was replaced twice per week. When the monolayer of adherent cells reached 80% to 90% confluence, cells were trypsinized using 0.25% trypsin-EDTA (Life Technologies) and subcultured to passage three.
Characterization of cells
Flow cytometry was used to characterize the surface marker expression of passage three ADSCs. Briefly, the medium was aspirated and the cell layer was washed with PBS before incubation with 1-mL 0.25% trypsin-EDTA for 3 minutes. Monoclonal antibodies against CD90, CD44, CD73, CD105, CD31, CD19, CD11b, and HLA-DR were used (all from BD Biosciences). Cells were analyzed using a FACSAria flow cytometer (BD Biosciences). Analysis was performed on 10,000 cells per sample and unstained cell samples were used to compensate for background autofluorescence levels.
Cell differentiation
For osteogenic induction, ADSCs were seeded on six-well plates at 3×105 cells/well. Next, the medium was replaced with osteogenic medium consisting of MEM α supplemented with 10% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin, L-glutamine, 0.1 µM dexamethasone, 50 µM ascorbate-2-phosphate, and 10 mM β-glycerophosphate (Sigma-Aldrich). The medium was changed twice weekly.
Alizarin red S staining
On day 21 of osteogenic induction, the medium was removed and cells were washed with PBS. Cells were fixed with 70% iced ethanol for 15 minutes at 4℃ and washed with distilled water. Alizarin red S staining solution was prepared by dissolving 1 g of Alizarin red S (Sigma-Aldrich) in 100 mL of distilled water, mixing, and adjusting the pH to 4.12 with 0.1% NH4OH. Images of Alizarin red S-stained cells were captured with a DS-U2 digital sight camera (Nikon, Tokyo, Japan).
Real-time polymerase chain reaction
Total RNA was extracted using TRIzol reagent (Life Technologies) according to the manufacturer's instructions. Each sample, containing 2 µg of RNA, was heated at 65℃ for 15 minutes, before addition of reverse transcriptase. cDNA was prepared through incubation at 50℃ for 60 minutes using the DiaStar RT Kit (SolGent, Seoul, Korea), and real-time polymerase chain reaction (PCR) was performed using a LightCycler instrument (Roche Applied Science, Indianapolis, IN, USA). LightCycler DNA Master SYBR-Green I (Roche Applied Science), cDNA template, primer pairs, and 25 mM MgCl2 were added to microcapillary tubes to give a final volume of 20 µL. PCR was conducted for 40 to 50 cycles, each consisting of predenaturation at 95℃ for 10 seconds, 5 seconds at a specific annealing temperature, and primer extension at 72℃ for 20 seconds. The expression level of the target gene was normalized to the β-actin expression level. Melting curves were visually inspected to confirm the specificity of product detection. The primer sequences were as follows: GLP-1r, TCAAGGTCAACGGCTTATTAG (forward) and TAACGTGTCCCTAGATGAACC (reverse); osteocalcin (OC), AGCAAAGGTGCAGCCTTTGT (forward) and GCGCCTGGGTCTCTTCACT (reverse); and alkaline phosphatase (ALP), CCCCCGTGGCAACTCTATCT (forward) and GATGGCAGTGAAGGGCTTCTT (reverse).
Western blot analysis
Cells were isolated using PRO-PREPTM Protein Extraction Solution (iNtRON Biotechnology, Seoul, Korea). Protein concentrations were determined using the BCA protein assay kit (iNtRON). GLP-1R protein was separated by 10% SDS-PAGE and electroblotted onto a Hybond-ECL nitrocellulose membrane (Amersham Biosciences, Little Chalfont, UK). The membrane was blocked with 5% skim milk and incubated with GLP-1R antibody (1:1,000 dilution; Abcam, Cambridge, UK), and subsequently with horseradish peroxidase-conjugated rabbit anti-mouse IgG (Cell Signaling Technology, Danvers, MA, USA). Immunoreactive bands were detected using the West-One Western Blot Detection System (iNtRON).
Statistical analyses
All statistical analyses were performed using SPSS version 17.0 (SPSS Inc., Chicago, IL, USA). For all variables, descriptive statistics, including the mean and standard deviation, were determined for each day. Relative GLP-1R gene expression levels were compared by repeated measures one-way analysis of variance to test for time-dependency. P<0.05 was considered to indicate statistical significance.
RESULTS
Flow cytometric surface marker expression analysis
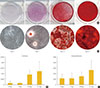
ADSCs exhibited fibroblast-like morphology and adhered to plastic (Fig. 1A). To confirm their multipotent differentiation potential, specific surface antigen expression was assessed according to the minimal criteria established by the International Society for Cellular Therapy [12]. The cells showed positive expression (≥95%) of mesenchymal stem cell (MSC)-specific antigen markers, including CD90, CD73, and CD105. Conversely, expression of CD11b (a hematopoietic cell marker prominently expressed on monocytes and macrophages), CD19 (a B cell marker), CD31 (a hematopoietic cell marker expressed on the surfaces of platelets, monocytes, and neutrophils), CD34 (a marker for primitive hematopoietic progenitors and endothelial cells), and HLA-DR was not detected (Fig. 1B).
Osteoblastic differentiation of ADSCs
Osteogenic differentiation was confirmed by cytochemical staining and gene expression analysis. After 21 days of osteogenic induction, positive Alizarin red S staining consistent with matrix calcification was observed (Fig. 2A). ALP is a commonly used marker for early osteogenic differentiation. In our study, ALP was notably upregulated at the 21 days (Fig. 2B). Expression of the late osteogenic marker OC is known to be upregulated immediately prior to mineralization, and we observed significant increases in OC mRNA levels by day 21 of differentiation (Fig. 2C).
Assessment of GLP-1R expression by Western blotting and PCR
Up-regulation of GLP1-R protein expression during the osteoblastic developmental sequence was confirmed by Western blot data for day 21 (Fig. 3A). Real-time PCR was used to assess the time-course of GLP-1R mRNA expression during osteogenic differentiation. For each time-point (0, 7, 14, and 21 days), the GLP-1R mRNA level of each sample was normalized to that of β-actin. The mean GLP-1R activity of ADSCs increased significantly in a time-dependent manner that was related to the degree of differentiation (P=0.013) (Fig. 3B).
DISCUSSION
In this study, we demonstrated that GLP-1R is expressed on ADSC-derived osteogenic cells, and that the level of expression increases in a time-dependent manner during differentiation. GLP-1 has been consistently reported to have a beneficial effect on bone in rodents [2,3]. Also, it is reported to have no adverse effects on bone, despite inducing weight loss in humans [13]. A GLP-1R agonist has been reported to prevent osteopenia by promoting bone formation and by suppressing bone resorption in aged, ovarectomised rats [14]. Furthermore, a GLP-1 agonist increased bone mineral density in type 2 diabetic rats [15]. The mechanism underlying this phenomenon is not clear. One possible explanation is that GLP-1Rs expressed on thyroid C cells promote calcitonin secretion, which inhibits bone resorption; however, GLP-1R was not detected in osteoblasts [16]. A second possibility is the presence of a functional receptor independent of the cAMP-linked GLP-1R. A final possibility is the existence of a GLP-1R.
The downstream effect of GLP-1 is mediated by a G-protein-coupled receptor that is expressed in various tissues, such as the pancreas, stomach, and vascular system [17]. Expression of GLP-1R by osteoblasts, however, has not been confirmed. GLP-1R has been reported to be expressed on thyroid C cells, which affect bone resorption by promoting calcitonin secretion, but the study did not detect GLP-1R expression in osteoblasts [16]. Conversely, expression of GLP-1R was reported in MC3T3-E1 cells, a well-known mouse osteoblastic cell line [4]. The authors suggested that this result indicates that, like other classic receptors, GLP-1R ha nd temperature-dependent. Another study reported that GLP-1R is expressed by osteoblastic cell lines, but that the expression level is cell line-dependent, perhaps reflecting different stages of osteoblast differentiation [5]. In that study, GLP-1R was expressed by young osteoblasts but not by mature osteoblasts derived from the Saos-2 osteosarcoma cell line. It was suggested that GLP-1R is expressed by osteoblasts, but that the expression decreases with maturation. However, GLP-1R expression was recently reported in osteocytes, which are derived from osteoblasts [15]. In addition, GLP-1R expression may differ among species [18,19].
We also evaluated the levels of OC and ALP in our ADSC samples. One of the most common methods used to examine mineralization is Alizarin red S staining, which indicates extracellular calcification. Alizarin red staining alone, however, may not be sufficient to confirm osteogenic differentiation [20,21]. ALP is a well-known indicator of the early stage of osteogenic differentiation of MSCs, and expression of ALP increases with osteoblast maturation [22]. In this study, ALP levels increased in a time-dependent manner during osteogenic differentiation of ADSCs. OC is a noncollagenous protein found in bone and dentin and is generally considered to be a relatively late-stage marker for the period of osteoblastic differentiation immediately prior to mineralization [23,24]. In our study, expression of both ALP and OC was elevated on day 21 of differentiation, confirming that osteogenic differentiation of ADSCs had occurred.
In summary, we demonstrated GLP-1R expression during osteogenic differentiation of ADSCs. Even though it is not yet clear whether GLP-1 has a role in bone metabolism, our results suggest that it may induce osteogenic differentiation in bone tissue. Further studies are needed to explore this possibility.

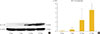
 XML Download
XML Download