

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Facioscapulohumeral muscular dystrophy type 1 (FSHD1) is a common muscular dystrophy characterized by an asymmetric descending progression starting in the face or shoulder-girdle muscles. FSHD1 is caused by contraction of the D4Z4 repeat array on chromosome 4q35 to a size of 1 to 10 units. The clinical severity of FSHD1 is inversely correlated with D4Z4 repeat units, and many patients with 7 to 10 D4Z4 repeat units have mild disease activity and remain nonpenetrant.12 Additionally, the D4Z4 repeat of 8−10 units is found in 1−3% of the healthy control populations.3 However, the repeat units are not always predictive for clinical severity, and patients with 8 to 10 D4Z4 repeat units occasionally revealed early symptom onset and severe clinical phenotype.1 Actually, the SMCHD1 and DNMT3B genes have recently been identified as genetic modifiers in FSHD1.45
The FAT1 gene is located on chromosome 4q35 proximal to the D4Z4 repeat array. The FAT1 mutations are linked to various diseases including schizophrenia, bipolar disorder, and cancer. It is known that this gene influences muscle patterning, regionalized muscle wasting, and adult muscle fiber functions.6 Furthermore, faulty regulation of the FAT1 expression was observed in FSHD1 muscle tissue, and mutations in FAT1 were found in patients with a FSHD-like phenotype.78
To identify the existence of a genetic modifier in Korean patients with FSHD1, we performed targeted sequencing of SMCHD1, DNMT3B, and FAT1 genes in patients with 8−10 D4Z4 repeat units. Then, we found that one patient had both 8 D4Z4 repeat units and a FAT1 mutation.
CASE REPORT
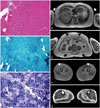
A 59-year-old woman in the MF1147 family presented to our neurologic clinic with proximal muscle weakness. She did not have a family history of any neuromuscular disorders. She first had gait disturbance and low back pain since her twenties. She felt weakness in shoulder girdle muscles at the age of 56 years. Then, she had had difficulty to rise from the floor since the age of 57 years. When we examined her at 59 years, she could walk independently. She displayed asymmetric and predominant weakness of shoulder and hip girdle muscles. Additionally, she had facial muscle weakness. She did not exhibit sensory deficits and joint contractures. Her serum creatine kinase level was 140 IU/L (reference value: <135 IU/L). Needle electromyography was compatible with generalized myopathy. A biceps brachii muscle biopsy was performed (Fig. 1A-C). Hematoxylin and eosin staining revealed increased fiber size variability, increased endomysial fibrosis, and multifocal degenerating fibers with lymphocytic infiltration. Modified Gomori-trichrome staining did not demonstrate any intracytoplasmic inclusions and subsarcolemmal depositions. Staining with reduced nicotinamide adenine dinucleotide-tetrazolium reductase showed mild disorganization of intermyofibrillar network. Whole body MR images were performed (Fig. 1D-G). At the upper arm level, T1-weighted MR images demonstrated predominant fatty replacement of right biceps brachii muscle. At the abdominal level, both abdominal and paraspinal muscles were found to have been totally replaced by fat tissues. At the thigh levels, semitendinosus, semimembranosus, biceps femoris, vastus medialis, and vastus intermedius muscles were shown to be totally replaced by the fat tissue. Rectus femoris muscle on the right side was severely affected. At the calf level, the MR image revealed bilateral soleus muscles predominantly affected.
Based on these clinical, pathological, and radiological presentation, we assumed that her disease was FSHD1. To determine the size of the D4Z4 repeat arrays, we performed Southern blot analysis and found 8 D4Z4 repeat units, therefore, she was diagnosed with FSHD1. Subsequently, we performed targeted-next generation sequencing of modifier genes including the SMCHD1, DNMT3B, and FAT1 genes to find the existence of a genetic modifier in Korean patients with FSHD1. Targeted sequencing was performed in 9 unrelated patients. All patients had asymmetric descending progression starting in the face or shoulder-girdle muscles and carried a contracted allele from 8 to 10 D4Z4 repeat units. Finally, we identified previously reported c.10331 A>G (p.N3444S) variant in FAT1 gene in the present case. This FAT1 variant was previously reported as a mutation in a myopathy patient who had a FSHD-like phenotype but did not have the contraction of the D4Z4
repeat units.8
DISCUSSION
In the present study, we identified a FAT1 mutation in a Korean patient with the shortening. The c.10331 A>G FAT1 variant has rarely been reported in dbSNP142 and the Exome Variant Server with allele frequencies of 0.00008. However, in vitro functional study demonstrated that this variant induced partial alteration of splicing by breaking an exonic splicing enhancer.8 Therefore, c.10331 A>G variant in the FAT1 gene is a pathogenic variant and influenced clinical phenotype of the present patient.
The FAT1 gene encodes a single pass transmembrane protein with the extracellular portion consisting of 34 cadherin repeats, five EGF-like domains and a laminin-G like domain. This protein belongs to the FAT cadherin superfamily and influences multiple biological activities, including smooth muscle cell motility, actin accumulation, cell polarity, and migrating muscle precursors.9101112 Recent studies demonstrated that the FAT1 expression is closely related with disease onset and the alterations in FAT1 gene cause FSHD-like myopathy.78 However, the pathogenic mechanism of FAT1 mutations in myopathy remains unclear.
The present case revealed facial diplegia, asymmetric descending progression, and lymphocytic infiltration. It was compatible with FSHD1, but we could not find any add-on effects of FAT1 alterations. It is probably because that the clinical presentation of FAT1 myopathy closely resembles that of FSHD1. Previous study demonstrated predominant involvement of facial, scapular, and humeral muscles and progression of the disease toward the lower limbs in patients with FAT1 mutations.8 These findings are typical clinical features of FSHD1. We initially planned neurological examination and segregation study of the D4Z4 contraction and the FAT1 mutation in the family. However, the patient and her family members refused further evaluation and genetic testing. Further studies are needed to identify the role of a genetic modifier of FAT1 gene in FSHD1.
In conclusion, our study is the first report of coexistence of the contracted D4Z4 repeats and FAT1 mutation in a FSHD1 patient.
 XML Download
XML Download