

 PDF
PDF ePub
ePub Citation
Citation Print
Print




INTRODUCTION
Helicobacter pylori is an important pathogen that causes many gastric diseases such as gastirits, gastroduodenal ulcers, and gastric cancers (1). The anti-H. pylori immunoglobulin G (IgG) seropositivity rate was lowest in infants aged 7–9 months among populations from neonates to 60–69 years and increased with age in Korea (2); the most primary H. pylori infections occur in early childhood (23). The clinical diagnosis of early H. pylori infection is difficult due to the nonspecific presentations of H. pylori infection such as abdominal pain and vomiting (4), and young children cannot explain the symptoms.
Early diagnosis of H. pylori infection in young children could be helpful for investigating the disease-associated pathogenesis, and identification of the proteins that induce the early human immune response would enable early diagnosis. The primary immune response involves immunoglobulin M (IgM) antibody, and secondary exposure to the same antigen produces IgG, immunoglobulin A (IgA), and/or IgE antibodies. After H. pylori infection, IgM antibodies against early antigens of H. pylori are formed and detected transiently in the acute period. Although IgM-based serological diagnosis of H. pylori infection was observed in early infants (5), most serological tests for H. pylori infection in children are based on IgG or IgA antibodies (267). However, the sensitivity of commercial enzyme immunoassays (EIAs) was low (22.0%–33.3%) in children aged 0–6 years and varied by commercial EIA (6). This low sensitivity of commercial EIAs suggested that young children might have a different immune response to H. pylori (6). In contrast, commercial EIAs testing IgG and IgA were suitable for serodiagnosis of H. pylori infection in children aged 1–14 years, and the most recognized bands were under 40 kDa (5).
One-dimensional (1D) immunoblots show only the molecular weight (MW) of antigenic proteins that react with IgG or IgA antibodies in sera. Anti-CagA positivity on 1D immumoblots correlated with the high anti-H. pylori IgG/IgA titers in the sera of H. pylori-positive children even younger than 5 years (9). In contrast, two-dimensional (2D) polyacrylamide gel electrophoresis (PAGE) separates proteins according to both isoelectric point and MW, thereby providing superior resolution of individual proteins from a complex mixture. 2D immunoblots identified CagA as the most reactive antigen recognized by all the H. pylori-positive sera (IgG) even in children less than 3 years old (8). Furthermore, protein excision from 2D gels often allows their analysis and identification through techniques such as peptide mass fingerprinting (10).
To our knowledge, no previous study investigated anti-H. pylori IgM positivity in the sera of H. pylori-positive infants. In the present study, we identified the operating antigens of H. pylori during early infection in infants by 2D-immunoblot analysis and analyzed the proteomics of H. pylori antigens that reacted with IgM antibodies in sera from H. pylori-infected infants.
MATERIALS AND METHODS
Subjects
From 1999 to 2005, infants diagnosed with H. pylori infection using histopathologic findings or urease tests and whose serum samples and gastric biopsy specimens were preserved in the Gyeongsang National University Hospital Biobank of Korea. H. pylori infection was confirmed by immunohistochemical detection using a polyclonal antibody and immunoelectron microscopic examination (Immuno-EM) of preserved gastric biopsy specimens as reported previously (11).
Bacterial strains
The strains were isolated in previous study (12) and deposited in the H. pylori Korean Type Culture Collection (HpKTCC, http://hpktcc.knrrc.or.kr/, Gyeongsang National University School of Medicine). Frozen H. pylori strain 51 was thawed and grown on Brucella agar plates containing 10% bovine serum, vancomycin (10 μg/mL), nalidixic acid (25 μg/mL), and amphotericin B (1 μg/mL) at 37 °C under 10% CO2 and 100% humidity. After an 18-h subculture, bacterial masses were collected to prepare protein specimens for 2D electrophoresis (2DE) analysis.
2DE and image analysis
1DE and 2DE samples were prepared as described previously (13). After washing with 40 mmol/L Tris-hydrochloride (HCl) (pH 7.2) and 1 mmol/L ethylenediaminetetraacetic acid (EDTA), bacterial pellets were lysed using a buffer containing 9.5 mol/L urea, 4% 3-((3-cholamidopropyl)dimethylammonium)-1-propanesulfonate (CHAPS), and 35 mmol/L Tris-HCl (pH 7.2). The solubilized protein samples (30 μg) were mixed with the rehydration solution containing 8 mol/L urea, 4% CHAPS, 10 mmol/L dithiothreitol (DTT), and 0.2% carrier ampholytes (pH 3.0–10.0) to a final volume of 125 μL and applied to immobilized pH gradient (IPG) strips (7 cm; Bio-Rad, Hercules, CA, USA) of pH 3.0–10.0 in a re-swelling tray (Bio-Rad). After the IPG strips were rehydrated, isoelectric focusing (IEF) was performed using a Protein IEF Cell (Bio-Rad), and 3 preset programs consisting of the first conditioning step (15 minutes, 250 V), the linear voltage ramping step (3 hours, 4,000 V), and the maximum voltage ramping step of up to 30,000 Vh. The current did not exceed 50 μA per strip. Following IEF, the strips were equilibrated with 0.375 mol/L Tris buffer (pH 8.8) containing 6 mol/L urea, 2% sodium dodecyl sulfate (SDS), 20% glycerol, 2% DTT, and 0.01% bromophenol blue. The strips were equilibrated again with the same buffer supplemented with 2.5% iodoacetamide. The 2D SDS-PAGE was carried out overnight at 20 mA per gel using a 12.5% separating polyacrylamide gel (8–10 cm) without a stacking gel. The resolved protein spots on the gels were visualized by silver staining (14) and scanned using a Fluor-S MultiImager (Bio-Rad). The spot intensities of each sample were determined and analyzed using PDQUEST 2D Gel Analysis Software Version 6 (Bio-Rad) installed on a Magic Station M5660 (Samsung, Seoul, Korea).
Immunoblot analysis
Protein bands were electrophoretically transferred from the SDS-PAGE gel onto a 0.2-μm nitrocellulose membrane using a semi-dry transblot (Bio-Rad) at a constant current of 120 mA for 60 minutes. After the transfer, unbound sites on the nitrocellulose membrane were blocked for 1 hour at room temperature with blocking solution (1% bovine serum albumin, BSA-TBST solution; Tribioscience Inc., Palo Alto, CA, USA) and washed 3 times in TBS with 0.05% Tween 20 (Alfa Aesar, Haverhill, MA, USA) for 5 minutes. Thereafter, the membrane was cut into strips, and each strip was incubated with individual serum samples diluted five-fold.
Immunodetection was performed with alkaline-conjugated monoclonal rabbit anti-human IgG (1:10,000), IgA (1:500), and IgM (1:500) antibodies for 30 minutes (Bethyl Laboratories Inc., Montgomery, TX, USA). Substrate development was performed using alkaline phosphatase buffer, 0.03% nitroblue tetrazolium (Thermo Fisher Scientific Inc., Rockford, IL, USA) and 0.015% 5-bromo-4-chloro-3-indolyl phosphate (BCIP) substrate (Thermo Fisher Scientific Inc.). The MW of antigenic proteins was measured using a protein standard (Bio-Rad).
Image analysis and in-gel protein digestion
A gel image was obtained by scanning the silver-stained gels with a Fluor-S Multimager (Bio-Rad) and documented with the PDQUEST 2-D Gel Analysis Software Version 6 (Bio-Rad) installed on the Magic Station M5660 (Samsung, Korea). Silver-stained spots were excised from 2DE gels and transferred into Eppendorf tubes. The chemical reducers mixture was prepared fresh with a 1:1 ratio of 30 mmol/L potassium ferricyanide and 100 mmol/L sodium thiosulfate. One hundred microliters of the mixture was added to the Eppendorf tube and occasionally vortexed until the brownish color disappeared. After the gel pieces were rinsed three times with distilled water (DW) to stop the reaction, 500 μL 200 mmol/L ammonium bicarbonate was added to cover the gel for 20 minutes and then discarded. The gel piece was dehydrated with 100 μL acetonitrile and dried in a vacuum centrifuge. In-gel digestion was carried out using the method described by O'Connell and Stults (14). Gel pieces containing protein spots were rehydrated by adding a digestion buffer on ice. The buffer containing 12.5 ng/mL trypsin was added to cover the pieces and incubated for 45 minutes on ice. The enzyme solution was removed and replaced with 20 μL buffer without enzyme, keeping the gel pieces wet during enzymatic cleavage overnight at 37°C, and the gel pieces were subjected to vigorous vortexing for 30 minutes. Twenty microliters digested solution was transferred into a clean Eppendorf tube and dried in a vacuum, and the resulting samples were dissolved in 2 μL 0.1% trifluoroacetic acid (TFA).
Peptide mass fingerprinting
A matrix solution composed of α-cyano-4-hydroxycinamic acid (40 mg/mL) in 50% acetonitrile and 0.1% TFA was prepared for peptide mass fingerprinting. The matrix solution and sample solution were mixed at a 1:1 ratio, and 2 µL was applied onto the matrix-assisted laser desorption/ionization time of flight (MALDI-TOF)/TOF target plate, rapidly dried, and washed using deionized water. The mixture solution was dried for 10 minutes at room temperature and subjected to MALDI-TOF-mass spectrometry (MS) and MS/MS analysis using an ABI 4800 Plus TOF-TOF Mass Spectrometer (Applied Biosystems, Framingham, MA, USA). The instrument was set at 200-Hz Nd:355-nm YAG laser operation. Peaks with signal/noise ratios > 25 were selected, and the 10 most intense ions were used for MS/MS analysis in 1 kV mode, 1,000–1,250 consecutive laser exposures.
All MALDI-TOF-MS spectra were searched against the National Center for Biotechnology Information (NCBI) protein database (version 20140415; 2,845 sequences, 921,323 residues) of Helicobacter 51 proteins with a molecular mass range ± 15% of that estimated from 2DE and allowing a peptide mass accuracy of 50 ppm. If matched proteins were absent, the molecular mass window was extended. MS/MS spectral data were analyzed using Protein Pilot V.3.0 database software (with the MASCOT V.2.3.02 database search engine) at 50 ppm mass tolerance. Pyroglutamic acid modification of N-terminal glutamine, oxidation of methionine, and acrylamide modification of cysteine were considered. Individual peptide ion scores were searched using a statistically significant threshold value of P = 0.050.
Ethics statement
All of the serum and gastric biopsy specimens in paraffin blocks that we experimented had been prepared and donated by the Gyeongsang National University Hospital, a member of the Korea Biobank Network after its Institutional Review Board reviewed the research protocols (GNUHIRB-2014-09-024). The board exempted informed consent because of its retrospective nature of research.
RESULTS
Subjects
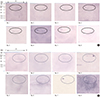
Eight infants were confirmed with H. pylori infection using urease tests, hematoxylin-eosin (H-E) staining, pre-embedding Immuno-EM, or immunohistochemistry (Table 1). The median age was 6.5 months (range, 4–11 months), and 2 were female. Upper gastrointestinal (UGI) bleeding was the most common clinical presentation. Urease test results showed all positive in body and seven positive in antrum. H-E staining revealed mild chronic gastritis in 7 infants, mild active gastritis in 1 infant, and no H. pylori infiltration in all. All 8 were confirmed with H. pylori infection by immunohistochemistry and pre-embedding immuno-EM (Fig. 1).
Table 1
Clinical data for infants enrolled in this study

Fig. 1
Histopathologic findings of the gastric antral mucosa in infant 4. (A) H-E-stained gastric mucosa (×400) showed a mild degree of chronic gastritis. (B) Immunohistochemistry with rabbit anti-H. pylori polyclonal antibody and DAB as a chromogen showed brown-colored gastric epithelial lining. (C) Immunoelectromicroscopic examination (× 12,000). Electron microscopic view of the immunohistochemical-stained area revealed several H. pylori.
DAB = 3,3′-diaminobenzidine-tetrahydrochloride-dihydrate

1D immunoblotting
1D immunoblots of IgG, IgA, and IgM antibodies for whole-cell proteins of H. pylori strain 51 were performed in 6 infants (Fig. 2). The sera of two infants were insufficient for both 1D and 2D immunoblots. IgG positivity to 120-kDa bands was observed in infants 1, 2, 3, and 5. IgA- and IgM-reactive bands were observed in infants 5, 6, and 9 and were around 60 kDa, 45 kDa, and under 30 kDA.
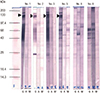
2D immunoblotting
2D immunoblotting with the IgG antibody showed reactive spots of approximately 120 kDa and pI 8–9 in 7 infants. 2D immunoblotting with the IgM antibody showed reactive spots around 43–68 kDa and pI 4–8 (Fig. 3A). The reactive spots of IgA antibodies were weaker and fewer than those of IgM antibodies (Fig. 3B).
Fig. 3
2D immunoblotting of 8 infant serum samples. (A) With IgM. Similar positive spots were observed in each membrane (circles). (B) With IgA. Similar and week positive spots were observed in each membrane (circles).
2D = two-dimensional, IgM = immunoglobulin M, IgA = immunoglobulin A.
Eighteen distinctive IgM-reactive spots from the 3-month-old infant 2 were numbered (Fig. 4) and compared with the 2DE pattern of whole-cell lysates of H. pylori strain 51 (Fig. 5). These 18 IgM-immunoreactive spots were excised for MALD-TOF-MS analysis. Ten peptide fingerprints were obtained and searched against the NCBI database (Table 2). Five proteins were identified: flagellin A (FlaA), urease β subunit (UreB), pyruvate ferredoxin oxidoreductase (POR) α subunit, hemolysin secretion protein in precursor (HlyB), and translation elongation factor Ts (EF-Ts).

Fig. 5
2D PAGE of whole-cell lysates of H. pylori strain 51.
2D = two-dimensional, PAGE = polyacrylamide gel electrophoresis.

Table 2
Early IgM-reactive H. pylori antigen proteins in infant serum samples

DISCUSSION
In the present study, H. pylori infection was confirmed in infants by immnuohistochemistry and immuno-EM, and serum samples were tested for the anti-H. pylori IgM antibodies to identify the antigens of early-stage H. pylori infection. The 18 IgM-reactive spots on 2D immunoblots were identified as H. pylori FlaA, UreB, POR, HylB, and EF-Ts by peptide mass fingerprinting using MALDI-TOF-MS.
Several proteomics studies reported that different antigens are recognized from different gastric diseases (151617) and suggested that this might owe to persistence of strong inflammation and tissue damage leading to increased antigen presentation during ulcer development. We identified 5 IgM-reactive proteins from infants less than 1 year old that were different from the translation elongation factor EF-G (FusA), catalase (KAtA), and urease α subunit (UreA) identified in duodenal ulcers (15) or GroES (also referred to as heat shock protein A) in gastric cancer (16). In addition, H-E staining revealed mild chronic gastritis in all infants and mild active gastritis in 1 infant.
We did not observe reactive spots against CagA in the 2D IgM immunoblots although CagA has been an immunodominant antigen in children (89). In 22 H. pylori-infected Japanese children aged 2–13 years, serum IgG distinguished reactive spots identified as CagA, CagA fragments, FlaA, Omp21/AlpB, and VacA on 2D immunoblots (8). The presence of CagA bands on 1D immunoblots correlated with the high titer of anti-H. pylori IgG antibody in children aged 0–15 years (9). IgG antibodies were observed on days 63 and 857 in twin boys with acute H. pylori infection (17). Serial 1D immunoblots of IgM antibodies, using 3-, 7-, and 9-month serum samples of an infant, showed 25, 29, and 47 kDa bands for the 3-month sample; a 120 kDa (CagA) band and several 60 kDa bands for the 7-month sample; and 120, 87, 61, 56, and 54 kDa bands for the 9-month sample (18). The absence of anti-CagA IgM at 3 months was considered as acute infection, and its presence at 7 and 9 months was considered as persistent infection (18). These results suggested that CagA induces gastric mucosal inflammation 2 months after initial H. pylori infection and might not play an important role for the bacterium's survival in the gastric mucosa during early infection. Herein, the CagA-positive on the 1D IgG immunoblot was probably due to placental acquisition of maternal anti-CagA IgG given that the positive infants were younger than 7 months.
The 18 IgM-reactive spots were 33–97 kDa, consistent with previous observation of low MW bands (31 kDa and 61 kDa) in 1D IgM and IgA immunoblots for children with no IgG-positive band (5). In H. pylori-positive Australian adults, highly IgA-immunoreactive proteins were 40–80 kDa on 2D immunoblots (19); the similar MWs of immunoreactive proteins might be related to the IgA antibody, which is made in the gastric mucosa upon direct exposure to H. pylori.
In the present study, IgM-positive spots were identified as FlaA, UreB, POR, HlyB, and EF-Ts. H. pylori can actively move through mucus using its flagella, which are virulence factors. H. pylori flagella consist of two main components, FlaA and FlaB, with the majority being FlaA (20). Although a study in Japan showed that FlaA reacted with IgG in sera of uninfected children and was nonspecifically reactive in sera of infected children (8), it might play an important role for moving in the gastric mucosa during early infection. H. pylori urease comprises two structural subunits, the 26.5-kDa UreA and 60.3-kDa UreB, and 5 accessory proteins (UreC, UreD, UreE, and UreF), and its activity increases under acidic conditions (pH < 6.5) (21). POR is a housekeeping enzyme involved in energy metabolism in bacteria by converting pyruvate to acetyl-CoA in microaerophilic conditions such as the human stomach (22). HlyB belongs to one of the largest membrane protein families, the ATP-binding cassette (ABC) transporters (23). IgM antibodies against UreB, POR, and HlyB might be made through activation of these proteins during early-stage infection for survival in the acidic stomach. EF-Ts is an interacting protein involved in extending the nascent polypeptide chain during the elongation stage of bacterial translation (24). The IgM-positive antibody to EF-Ts suggested that active protein synthesis occurs in early infection.
It is important to note the following limitations in our study. First, we did not compare our patients to infants without H. pylori infection. The identified proteins were the antigens isolated from H. pylori on the NCBI database. Further studies will be needed to determine whether the identified proteins are associated with other diseases in children without H. pylori infection. In addition, we compared the results with those for H. pylori strain 51, which was isolated from a patient in our hospital, but identified the proteins using NCBI database entries from H. pylori strain J99 or 26695. H. pylori is believed to exhibit a large degree of genomic and allelic diversity (25). We did not compare the genomic sequences of H. pylori strains 51, J99, and 26695.
In summary, this is first study to evaluate the early immune response to H. pylori in infants utilizing proteomics. We identified 5 protein candidates for the essential and active proteins enabling survival and colonization in human stomach during early H. pylori infection. The identification of early immunogenic H. pylori proteins is a prerequisite for the development of serological test kits based on recombinant antigens and will support efforts for early diagnosis in infants.
In conclusion, IgM antibodies of 8 infant patients with H. pylori infection reacted with H. pylori proteins. Immunoblot spots were compared with those from whole-cell extracts of H. pylori strain 51, and 18 spots were excised, digested in gel, and analyzed by MALDI-TOF-MS. Of the ten peptide fingerprints obtained, the H. pylori proteins FlaA, UreB, POR, and EF-Ts were identified and appeared to be active during the early infection periods. These results might aid identification of serological markers for the serodiagnosis of early H. pylori infection in infants.
 XML Download
XML Download