

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Staphylococcus aureus (S. aureus) is one of the common causes of subclinical mastitis worldwide, which is of economic importance to the dairy industry [13]. Raw milk is a potential source of S. aureus in milk and milk products, especially in the case of defective pasteurization. The main reservoir of S. aureus seems to be the infected quarter. Molecular epidemiological analysis of the bovine S. aureus population suggested that a small number of clonal types were responsible for most infections, and that strains had a broad geographic distribution [7,11,15].
S. aureus has a capacity to produce a large number of putative virulence factors [6,8,9]. Some of these factors may be of more importance than others in different diseases or at different stages of the pathogenesis of particular infections, as not all factors are produced by each strain. At present, nothing has been reported about the occurrence of these virulence factors among S. aureus isolates from India, and about the possible distribution of single S. aureus clones as causative agents of bovine mastitis at various farms. The present study was conducted to genotypically characterize S. aureus isolates in milk samples from cows with subclinical mastitis.
Materials and Methods
Samples
A total of 552 quarter milk samples were collected from 140 cows selected randomly from 8 farms in the Vidarbha region of Central India. Samples were collected over a two month period. The samples were tested by the California mastitis test (CMT) for subclinical mastitis, and were graded as negative, trace, weak, distinct, or strongly positive [14]. Isolation of Staphylococcus was attempted from the CMT positive milk samples.
Genotypic characterization
Chromosomal DNA of the isolates was prepared as described by Wilson [19] with some modification. In brief, bacteria were grown in brain heart infusion broth for 24 h at 37℃. The cultures were centrifuged at 4℃ at 8,000 g for 10 min. The pellet was suspended in TE buffer (200 µl) (10 mM Tris HCl + 1 mM EDTA, pH 8.0) with lysozyme (10 mg/ml) and incubated at 37℃ for 2 h. The bacteria was lysed with 10% SDS and proteinase K (10 mg/ml), and was incubated at 65℃ for 30 min. The denatured protein, cell wall debris, and polysaccharides were eliminated by the addition of 5M NaCl and CTAB/NaCl (10% hexadecyl trimethyl ammonium bromide in 0.7M NaCl) and incubated for 30 min at 65℃. DNA was purified by extraction with phenol: chloroform (1 : 1) and chloroform: isoamyl alcohol (24 : 1). DNA was precipitated with isopropanol and sodium acetate (3 M) solutions, washed in 70% ethanol, and suspended in 50 µl of TE buffer.
The virulence determinants investigated using the oligonucleotide primers included the genes encoding coagulase (coa), clumping factor (clfA), the IgG-binding region and the X-region of protein A (spa), enterotoxin A (entA), and thermonuclease (nuc). For all the genes, reaction mixtures (25 µl) included 2 µl template DNA, 10 × PCR buffer (Sigma Aldrich, USA), 25 mM MgCl2, 200 µM of the four dNTPs, 10 pmol of each of the 2 primers (Bangalore Genei, India), and 1U Taq DNA polymerase (Sigma Aldrich, USA).
In the present study, the amplification parameters and primer sequences described by Straub et al. [18] were used (Table 1). The amplification of genes was carried out with thermocycler (Thermo Hybaid, USA).
Amplified products were separated by agarose gel electrophoresis (1.5% agarose containing 0.5 mg ethidium bromide in 0.5 × Tris-EDTA electrophoresis buffer) at 5 V/cm for 2 h and photographed under UV illumination.
Results
Isolation of Staphylococcus was attempted from CMT positive milk samples. Out of 552 milk samples collected from 140 cows on 8 farms, 501 (90.76%) samples from 134 cows were found to be CMT positive. Of these 268 milk samples, 114 cows harbored Staphylococcus sp. On the basis of cultural and biochemical properties, 37 isolates were identified as S. aureus. All 37 isolates were positive for the tube coagulase test. Others strains were identified as S. intermedius, S. hyicus, coagulase negative staphylococci, and Micrococcus (data not shown).
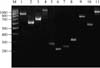
Amplification of the coa gene yielded three different products of 627, 710, and 910 bp for 20, 10, and 7 isolates from 7, 4, and 5 farms, respectively, and gene polymorphism was observed in isolates originating from 5 farms. All of the isolates yielded an amplicon with a size of approximately 1,042 bp of the clfA gene. The amplification of the gene segment encoding the IgG binding region of protein A (spa) revealed a size of 590, 810, and 970 bp in 12, 15, and 7 isolates from 5, 5, and 4 farms, respectively, and gene polymorphism was noted in isolates from 4 farms. The X-region binding of the spa gene produced an amplicon of 220, 253, and 315 bp in 10, 9, and 7 isolates, respectively. The amplification of the extracellular thermonuclease nuc gene produced an amplicon of 279 bp in 36 out of 37 isolates. All of the samples were found to be negative for the entA gene. Amplicons specific to the coa, clfA, nuc, spa IgG binding, and X-region genes are shown in Fig. 1. The genotypic properties of the 37 S. aureus isolates are summarized in Table 2.
Discussion
S. aureus has been recognized as a pathogen in human and animal infections. In the present study, 37 S. aureus strains isolated from subclinical bovine mastitis cases were identified and further characterized by PCR amplification of various virulence genes encoding clumping factor and coagulase activity, and gene segments encoding the immunoglobulin G-binding region and X-region of protein A and stable thermonuclease activity. Comparable PCR-based detection studies of the virulence genes have been described by other investigators [1,15].
The coa and spa (IgG-binding region and X-region) genes investigated in the present work exhibited typical gene polymorphism. This attribute could be used for the genotypic characterization of single isolates of this species. The spa gene segments encoding the X-repetitive region are known to consist of a variable number of small repeats [5]. It is thought that the spa domain encoding the X-region may serve to extend the N-terminal IgG-binding portion of the protein through the cell wall. It was interesting to note that isolates from the same farm exhibited polymorphism among the coa and spa genes.
The ability of S. aureus to adhere to extracellular matrix proteins is thought to be essential for the colonization and the establishment of infections [5]. S. aureus possesses various adhesion genes, including clfA, fnbA, and cna [16]. PCR analysis of the other virulence genes revealed the nuc and clfA genes in 36 and 37 strains, respectively, of the 37 strains investigated, suggesting an important role of these elements in the pathogenecity of bovine mastitis. However, entA was not present among the strains. In contrast, combined occurrence of enterotoxin genes has been described by other investigators [1,10,17,20].
In the present study, S. aureus isolates from cattle with bovine mastitis were found to differ in their gene patterns. Phenotypic and genotypic characterization might provide a better understanding of the distribution of the prevalent S. aureus clones among bovine mastitis isolates. This can aid in the investigation and control of S. aureus infections in dairy herds.


 XML Download
XML Download