

 PDF
PDF ePub
ePub Citation
Citation Print
Print


The physiological control of feeding is one of many behavioral functions that are influenced by the hypothalamic-pituitary gonadal (HPG) axis [1]. For survival and reproductive success, reproductive hormones have co-adapted to modulate ingestive behavior, energy storage, and energy expenditure [2]. Hormones have multiple effects on food intake, body weight, carcass composition, and voluntary exercise in rats, suggesting that hormone-induced behavior changes may alter body weight and levels of adipose tissue [3,4]. Another study suggested that variations in the estrous cycle may affect meal size and body weight in female rats [5].
Leptin, a protein secreted from adipose tissue, has been proposed to be a satiety factor that reduces food intake and enhances energy expenditure, leading to reduced fat storage [6,7]. Plasma leptin levels are directly proportional to existing fat reserves. Leptin not only controls food intake, but it is also involved in reproductive neuroendocrine functions, which are controlled by the hypothalamus [8]. Leptin communicates the sufficiency of energy stores for reproduction by promoting the activity of GnRH neurons [9]. The response to leptin changes during hormonally induced estrous cycles in rats may cause cyclic feeding behavior in rats [1]. While food intake and body weight of rats fluctuates during the estrous cycle, it remains to be determined whether changes in leptin responses occur during the rat estrous cycle. The present study was conducted to determine the concentration of serum leptin and the expression of leptin mRNA in adipose tissue as well as food intake during the estrous cycle of intact female rats.
Materials and Methods
Animals
Twenty-four specific-pathogen-free Iar: Wistar-Imamichi female rats were obtained from the Imamichi Institute of Animal Reproduction (Tsuchiura, Ibaraki, Japan) at 7 weeks of age. The animals were seronegative for Mycoplasma pulmonis, Bacillus piliformis, Boldetella bronchiseotia, Streptococcus pneumoniae, and Sendai virus. They were housed in individual cages at a temperature of 24±2℃ in a light-controlled room (12:12 h light:dark cycle). Food (Oriental MF: Oriental Yeast Co., Ltd., Tokyo, Japan) and tap water were provided ad libitum. The animals were allowed to acclimatize for seven days after arrival to the animal facilities. The animals were around 197±4.2 g and 9 weeks of age, when entering the study. The animal studies were approved by the Institutional Animal Care and Use Committee of the Nippon Veterinary and Life Science University and carried out in accordance with the National Institutes of Health, USA Guide for the Care and Use of Laboratory Animals. All efforts were undertaken according to the "3R principles" of the directive to reduce the number of animals used in this study, and optimize experimental protocols for obtaining maximum data from each tested animals.
Vaginal smear
On reaching the age of 8 weeks, estrous cycles were monitored daily using vaginal smears taken between 14:00 and 15:00 h. Smears were examined under a light microscope to estimate the relative preponderance of cornified epithelial cells, nucleated epithelial cells, and leukocytes [10]. Female rats that had experienced at least two consecutive 4-day estrous cycles were used for these studies.
Measurement of diet and leptin
Food consumption was measured by weighing the food hoppers at 07:00 and 19:00 h; a visual check was made for any diet dropped on the floor of the cage. Measurements were taken over the period of the 4-day estrous cycle.
Rats in proestrous and diestrous were decapitated between 14:00-15:00 and 19:00-20:00 h, and blood samples were collected from the vena cava cuadalis and centrifuged at 2,200 g for 15 min at 4℃. The serum was stored at -35℃ until it was analyzed for leptin concentration using a commercially available ELISA Kit (Rat Leptin ELISA KIT(U-type) AKRLP-D10; Shibayagi Co., Ltd., Gunma, Japan)
Leptin mRNA expression
For mRNA studies, visceral abdominal fat was dissected and immediately frozen in liquid nitrogen and stored at -80℃ until RNA extraction. Total RNA was isolated using an RNeasy Lipid Tissue Midi kit (Qiagen, Maryland, USA) according to the manufacturer's instructions. RNA samples were quantified spectro-photometrically (Nano drop, ND-1000, V3.12, Houston, TX, USA), and the integrity was confirmed using a 1.5% agarose gel containing 2.2 M formaldehyde. Total RNA was transcribed into first strand cDNA for each sample using a SuperScript™ III Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA) in a final volume of 20 µL; 1 µg RNA sample was added with 1 µL of 20 µM oligo dT Primer, and DNase- and RNase-free water. Samples were incubated at 65℃ for 10 min and then placed on ice. Then, 4 µL of 5x first-stand buffer, 2 µL of 0.1M dithiothreitol (DTT), and 4 µL of 10 mM 2'-deoxy-nucleoside 5'-triphosphate mixed solution (dNTP) was added. Samples were pre-incubated at 42℃ for 2 min, and then 0.5 µL of SuperScript™ III RT was added. Samples were incubated at 42℃ for 1 h, 30 µL of TE buffer (Tris-hydrochloride buffer, pH 8.0, containing 1.0 mM EDTA) was added, and the samples were heated at 65℃ for 10 min to inactivate the RTase.
Real time RT-PCR was performed using a specific forward primer (5'-GACCAGACCCTGGCAGTCTA-3') and reverse primer (5'-CTCAGCATTCAGGGCTAAGG-3') to amplify the product. Samples were normalized using the housekeeping gene glyceraldehydes-3-phosphate dehydrogenase (GAPDH) using a forward primer (5'-TGTCAGCAATGCATCCTGCA-3') and reverse primer (5'-CCGTTCAGCTCTGGGATGAC-3'). Real time RT-PCR was performed in a 25-µL final reaction volume using Platinum SYBR Green qPCR Supermix-UDG (Invitrogen, Carlsbad, CA, USA). Each real time RT-PCR contained 12.5 µL of Platinum SYBR Green qPCR Supermix-UDG, 10 µM of each primer, and an RNA template. Real-time RT-PCR was carried out in duplicate for each sample using an ABI 7500 Real-time RT-PCR System (Applied Biosystems, Foster City, CA, USA) with the following parameters: one cycle of 50℃ for 2 min (UDG incubation); 95℃ for 2 min; 40 cycles of 95℃ for 15 sec and 60℃ for 30 sec. Analysis of melting curves were performed with each series to confirm the specificity of the amplified products. To confirm the absence of genomic DNA signal, amplifications without reverse transcriptase were performed. Standard curves were prepared for tested and housekeeping genes. All expression data were normalized by dividing the amount of GAPDH used for the target gene by the amount used for the control.
Results
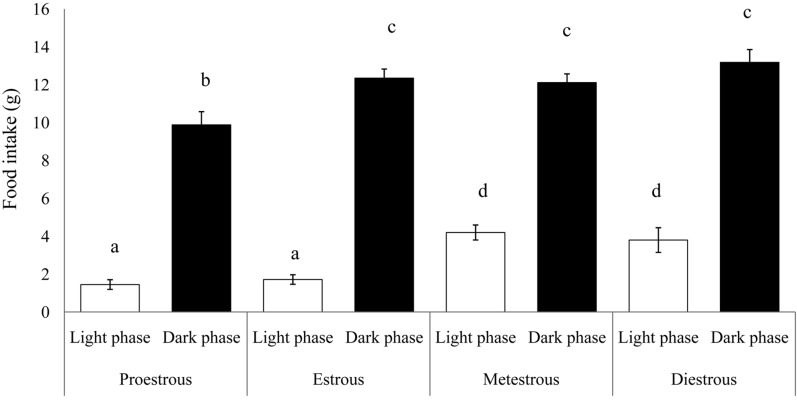
Food intake in intact female rats was measured over 4 days of the estrous cycle (Figure 1). Food intake was always greater (P<0.01) in the dark phase compared with the light phase, in all phases of the estrous cycle. Food intake in the dark phase in proestrous rats, however, was less (9.88±0.69 g) than that in the dark phase during the remaining of the estrous cycle (P<0.05). During the light phase, food consumption while in the proestrous or estrous phase, was lower than during the metestrous and diestrous periods (1.45±0.25 and 1.71±0.25 g, respectively) (P<0.01).
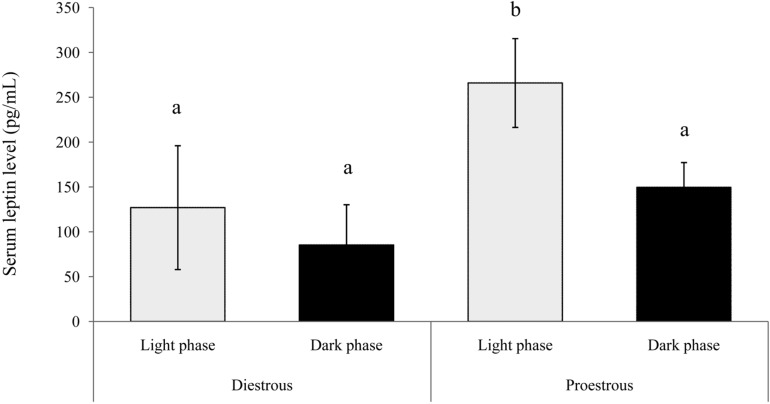
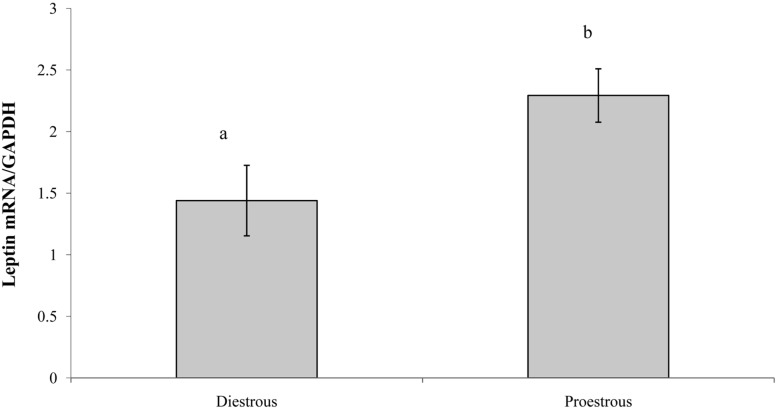
The concentration of serum leptin in proestrous and diestrous rats is presented in Figure 2. Serum leptin concentration was significantly higher (P<0.01) in the light phase of proestrous rats. The concentrations of serum leptin during the light and dark phases in diestrous rats were similar to that in the dark phase in proestrous rats. Similarly, leptin mRNA expression was higher in adipose tissue during the proestrous period (P<0.01) compared with the diestrous period (2.29±0.22 and 1.44±0.28 Leptin mRNA/GAPDH, respectively) (Figure 3).
Discussion
The present study demonstrates that food intake in intact female rats is higher in the dark phase compared with the light phase during the normal estrous cycle. Our results are in agreement with previous findings, which demonstrated that food intake in rats occurs mainly during the dark phase of the diurnal cycle [11-13]. Our observation that decreased food intake occurs during the light and dark phases of proestrous is similar to previous results showing that food intake is greatly reduced during proestrous [12]. However, Brobeck et al. [14] reported that decreased food intake occurred during the estrous phase. This pattern is a result of the estrogen surge that occurs around the time of increased physical activity and decreased eating activity, whereas estrogen levels, which alter thresholds of hypothalamic sensitivity, peak in the early morning of proestrous [12,14]. Decrease in food intake was facilitated by decreasing meal size due to a direct influence of estradiol on the CNS appetite centers, affecting the mechanism that terminates short-term food intake. These are not secondary to changes in the level of total daily food intake [15]. In the present study, serum leptin levels in proestrous and diestrous rats were high in the light phase compared with the dark phase. The levels were found in the light phase more highly in proestrous than diestrous rats. Leptin mRNA expression in adipose tissue was also higher in proestrous compared with diestrous rats. These results suggest that serum leptin levels are positively regulated by estrogen, as serum leptin concentrations peaked during periods of elevated estrogen activity in normal cycles. In contrast, serum leptin levels increased during the dark phase [13]. Bennet et al. [16] and Tanaka et al. [17] demonstrated that serum leptin levels increase during proestrous in rats. Evaluation of the body kinetics of leptin show that its half-life is only 9.4 min, indicating that the production rate varies significantly depending on hormonal factors such as the serum estrogen levels [18]. Other reports showed that serum leptin concentrations do not change significantly during the estrous cycle in rats [19-21]. Although some studies reported that estrogen stimulates the concentration of serum leptin and leptin mRNA expression in rat adipose tissue, which is mediated by the adipocyte estrogen receptor [1,17,22,23]. Sjoholm et al. [1] reported that hormonally induced changes in the responsiveness of leptin during the estrous cycle, particularly during the proestrous and estrous periods, and the mechanism of estrogen regulation of food intake involve an upregulation of leptin-receptor abundance and leptin sensitivity.
Leptin, which has been implicated in the regulation of reproduction, could act at multiple levels within the reproductive axis [9]. It prevents disruption of the estrous cycle in females, but only minimally restores LH level in males [24]. Leptin regulation of the reproductive system is mediated by its hypothalamic receptor eliciting the release of GnRH, which subsequently induces the synthesis and release of FSH and LH [9]. Tanaka et al. [17] reported that the elevation of serum leptin in the proestrous period might mediate the estrogen action that triggers preovulatory LH secretion. The LH surge is necessary for ovulation, which can induce estrogen levels during proestrous period.
In conclusion, our study show that food intake changes during the estrous cycle, and serum leptin concentration and leptin mRNA expression levels peak during proestrous in virgin female rats. Our data suggest that changes in serum leptin concentration during the estrous cycle may regulate cyclic feeding behavior in rats.
 XML Download
XML Download