

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
After the discovery of leptin in the mid-1990s, the role of adipose tissue had begun to gain attention, in that its role could include not only the storage of triglycerides but also the secretion of metabolic proteins.1 Leptin, now known to be released from adipose tissue, regulates the appetite, the balance of energy, and endocrine function, as well as the immune system.2 Other than leptin, adipokines such as adiponectin, resistin, and retinol binding protein-4 are secreted from adipose tissue, affecting insulin resistance.3-5 Furthermore, several studies have suggested that tumor necrosis factor α (TNFα) and interleukin-6, one of the most important cytokines of inflammation, are associated with adipocytes.6,7 In one study, it has been observed that visceral fat tissue promotes systemic inflammation by secreting cytokines into portal circulation.8 This systemic inflammation has an effect on insulin signaling and causes insulin resistance.9
Several receptors and metabolic enzymes for sex hormones mediate metabolism and inactivation of the hormones through their interaction in adipose tissue. Therefore, the concentrations of these circulating sex hormones are influenced by the amount and distribution of the adipose tissue.10 Evidence from a previous study showed that an increased level of testosterone in the bloodstream had caused a rise in insulin sensitivity and a decline in the risk of metabolic syndrome.11 In addition, testosterone was identified to have a negative correlation with obesity, and reciprocally, androgen was found to have an influence on the amount and the distribution of fat.12
This article describes the metabolism of androgen in adipose tissue based on the findings of recent research, and it reviews the effects of androgen on the distribution and metabolism of adipose tissue.
ADIPOSE TISSUE AND TESTICULAR FUNCTION
For the past fifty years, it was assumed that the substance responsible for suppression of food intake according to the increase in the stores of energy would exist in the brain.13 This substance had been discovered after homozygous mutation disrupting genes, ob, related to obesity, and db, related to diabetes, were identified in mice.14 An additional study reported that the ob locus had synthesized a substance associated with satiety while the db locus was identified as the crucial gene for this substance to take effect.15,16 This substance is termed leptin, and it was observed that the concentration of leptin increased in the bloodstream as the quantity of body fat increased.17 Another experiment on ob/ob mice with the inability to synthesize leptin and db/db mice with malfunctioning of leptin receptors indicated that both groups of mice showed less movement, less energy expenditure, and increased food intake, leading to obesity and infertility, known to be caused, in this case, by malfunctioning of spermatogenesis in the testes.18,19 When leptin was injected into these mice, only the ob/ob group recovered from infertility.20 These findings imply that obesity is not the sole cause of infertility, but rather, leptin plays a critical role in normal reproduction.
Spermatic cells and Leydig cells in the testes express receptors for leptin.21 This indicates that leptin may play a role in secretion of testosterone and in reproduction. Several lines of evidence have shown that leptin was involved in gonadotropin-stimulated testicular steroidogenesis.22-24 Moreover, an experimental study on mice in vitro suggested that when the anterior pituitary was treated with leptin followed by incubation, the concentrations of luteinizing hormone (LH) and follicle stimulating hormone had increased in proportion to the increase of leptin concentration; it also showed dose-responsiveness with an increase in LH-releasing hormone when the median eminence-arcuate nucleus was treated with low concentrations of 10-12~10-10 M leptin.24 To verify a similar effect of leptin in vivo, leptin was injected into the third ventricle of the ovariectomized estrogen-primed rat; it was found that the LH level increased as expected.24
Leptin also participates in spermatogenesis of the testis. According to research on the distribution of leptin receptors in the testes of the mouse, leptin had an effect on proliferation and differentiation of germ cells through phosphorylation of signal transducer and activator of transcription-3.25 Another study that compared a group of normal mice with a group of leptin-deficient ob/ob mice showed that impaired spermatogenesis, increased germ cell apoptosis, and up-regulated expression of proapoptotic genes were associated with leptin deficiency.26
ANDROGEN METABOLISM IN ADIPOSE TISSUE
About 50% of testosterone in adult males is bound to albumin, 44% is bound to sex hormone binding globulin (SHBG), and only 2~3% stays free-form.27 In fact, SHBG is known to decrease the metabolic clearance rate of testosterone and interrupts its movement into cells;28,29 thus, bioavailable testosterone from the tissues is in the unbound as well as the albumin-bound form.
Testosterone can take effect itself or after being transformed by 5α-reductase into the more powerful androgen dihydrotestosterone (DHT). Some of the testosterone is converted to estrogens by aromatase (ARO).30,31 On the other hand, another study discovered that ARO was present in the stromal cells of adipocytes in humans,32 indicating that adipose tissue was involved in the metabolism of sex hormones. Furthermore, increased ARO activity in obese males led to more androgens converting to estrogens, resulting in a higher level of estrogen and a decline of androgen in the plasma.33
It is well established that obesity has an impact on the hypothalamus-pituitary-testicular axis; a study on obese males showed that the Body Mass Index had a negative correlation with the concentration of testosterone and a positive correlation with estradiol.34 The same study indicated that the LH pulse frequency was similar in both normal and obese groups, while the mean diurnal LH levels, mean diurnal LH pulse amplitude, and the sum of all diurnal LH pulse amplitudes and secretory masses were noticeably lower in the obese group.34 Furthermore, an increase in pro-inflammatory cytokines such as TNFα from adipose tissue influences the secretion of gonadotropin in the pituitary.35
DHT, known as a potent androgen, does not get metabolized by ARO, but it has been observed that DHT is converted to the inactive metabolites 5α-androstane-3α and 17β-diol (3α-diol) in fat tissue.36,37 Aldo-keto-reductase 1C (AKR1c), which metabolizes DHT into its inactive form, is expressed in fat tissue.38,39 Evidence from an experiment on human adipose tissue demonstrated that AKR1C mRNA expression and DHT inactivation occurred in subcutaneous and visceral adipose tissue; also, mRNA levels were positively correlated with the amount of visceral adipocytes.37 Furthermore, the androgen inactivation rates were higher in the obese group than the normal group.37 In short, AKR1C mRNA expression increases with the mass of adipose tissue, leading to an increase in androgen inactivation rates.
THE EFFECTS OF ANDROGEN ON THE ADIPOSE TISSUE
The receptors for androgen are present in adipose tissue, and are more prominent in visceral (VAT) than subcutaneous adipose tissue (SCAT).40 Among various hormones that regulate the metabolism of adipocytes, catecholamine plays an important role in controlling lipolysis and receptors for catecholamines, known as adrenoreceptors, increase in density by testosterone. Testosterone also influences adipose tissue by up-regulating the adrenoreceptor (AR) that activates lipolysis.41 An experiment on adipose precursor cells of male rats suggested that testosterone stimulated catecholamine-induced lipolysis in a dose-dependent manner through β-AR and adenylate cyclase.42 In particular, lipolysis induced by testosterone was consistently observed, even after treatment with ARO inhibitor, proving that the effect was not due to the estrogens converted by ARO.42 An experiment on male rats in vivo also revealed that androgen had caused a decline in lipolysis through decreasing triglyceride/fatty acid cycling in adipose tissue when the androgen antagonist, cyproterone acetate, was injected into the rats.43 Furthermore, cyproterone acetate reduced catecholamine-induced lipolysis without affecting the activity of esterification enzymes such as GPAT or PPH that were required for the synthesis of triglyceride.43
Lipo-protein lipase (LPL) hydrolyzes circulating triglyceride-rich lipoproteins to facilitate the adipocyte activity of intracellular esterification that utilizes free fatty acids during the process.44 LPL is known to be associated with not only obesity but also with the energy balance, insulin action, and metabolic disorders such as abnormal body weight regulation.45 It is recognized that androgen reduces the LPL activity in adipose tissue, adjusting the central fat accumulation;46 in fact, an experiment that examined the effect of androgen on abdominal adipose tissue suggested that the LPL activity in the tissue had decreased and the triglyceride uptake was inhibited in the group in which testosterone was injected for nine months.47 Such an uptake of triglyceride was more clearly observed in VAT than SCAT, possibly due to the greater number of androgen receptors expressed in VAT.48
CONCLUSION
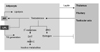
Leptin, produced in adipose tissue, influences the functioning of the testes directly and indirectly. The secretion of testosterone is directly mediated by the leptin receptors that are present in the leydig cells of the testes. Moreover, leptin acts on the hypothalamus-pituitary-testicular axis, regulating the functioning of the testes indirectly.
Adipose tissue is involved in the metabolism of androgen to estrogen known to be converted by ARO found in fat tissue. The adipose tissue also affects hypothalamus-pituitary-testicular axis, reducing the release of gonadotropin. A strong androgen DHT, in addition, is converted to an inactive metabolite by AKRIC present in adipose tissue.
Androgens have a negative effect on adipogenesis and lipid synthesis by up-regulating the AR for catecholamine, thus increasing lipolysis. In addition, androgens reduce the LPL activity that is necessary for intracellular esterification of adipocytes, adjusting central fat accumulation.
In conclusion, adipose tissue and androgen influence each other in a bidirectional and reciprocal way (Fig. 1).
 XML Download
XML Download