

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Osteoporosis, which could exacerbate after menopause, is a serious problem for elderly women worldwide [1]. It is widely accepted that 17-beta-estradiol (E2) deficiency plays a key role in the pathogenesis of post-menopausal osteoporosis (PMO) [2]. Indeed, estrogen can modulate bone production and bone resorption, exerting distinct effects on osteoblasts and osteoclasts directly via estrogen receptor (ER)–mediated mechanisms [34]. It was recently reported that estrogen inhibited the production of bone resorption-related cytokines such as tissue necrosis factor-α, granulocyte macrophage colony-stimulating factor (M-CSF), and prostaglandin-E2; stimulated osteoblast proliferation; and suppressed osteoclast activity [5]. Based on the anabolic actions of estrogen on bone remodeling, estrogen replacement therapy (ERT) has been used as a therapeutic modality in elderly women for the prevention or treatment of PMO [6]. However, the economic burden and risk of malignant changes in estrogen-sensitive organs, e.g., the breast, uterus, and ovaries, owing to long-term ERT, have strongly restricted its clinical applicability [78]. Furthermore, bone mineral density may decline even more sharply after the withdrawal of short-term ERT, increasing the risk of fracture. Thus, there is an urgent need for the development of alternative therapeutic options for PMO with high cost-effectiveness and no harmful effects.
Interest in the use of plant-derived estrogens, also known as phytoestrogens, has increased significantly. Phytoestrogens are natural polyphenolic compounds with structural similarity to E2 [9]. Although efficacies of phytoestrogens are not as high as those of endogenous estrogens, many patients with PMO self-prescribe these compounds and they have gained worldwide acceptance as beneficial and safe treatments. Based on their functional similarity with E2, most flavonoids, including isoflavones, isoflavanones, flavones, chalcones, and flavanones, are considered to be phytoestrogens [1011].
Many natural plants are known to contain a variety of flavonoids. Rhus verniciflua Stokes (RVS) is an Asian tree species of the genus Toxicodendron, native to China and the Indian subcontinent, and cultivated in regions of China, Korea, and Japan [12]. Commonly known as the lacquer tree, RVS has been utilized as a natural dye, food additive, and herbal medicine [13]. Although Dongui-Bogam (the Korean traditional medicinal encyclopedia) reported that RVS has beneficial health-promoting effects, including prevention of circulatory problems, improvement of blood homeostasis, antibacterial activity, and stimulation of the bone marrow function, the clinical therapeutic use of RVS has been restricted because of the presence of urushiol, an allergenic constituent, which might cause anaphylaxis [1415]. RVS has been reported to exert various biological activities, including anti-inflammatory, anti-allergic, antitumor, neuroprotective, and antithrombotic effects [16171819]. A recent study revealed that butein and sulfuretin, the flavonoid components of RVS, have strong affinities for the ER and consequently exhibit prominent estrogenic activity [20].
Based on the accumulated evidence of the various biological actions of RVS, including the estrogenic activity described above, we hypothesized that the long-term intake of RVS might prevent or delay the progression of PMO. To date, no reports have been published on the possible in vivo effects of RVS in relation to the prevention of PMO. To address this issue, this study evaluated the anti-osteoporotic activity of RVS in an experimental animal model of PMO, which was produced by ovariectomy of female Sprague Dawley (SD) rats. After the intragastric administration of RVS, we investigated the serological changes in markers that reflect the severity of PMO and the histologic changes in the uterine tubes and the epiphyseal plates in the tibia. Furthermore, using microcomputed tomography (µCT), various indices that were indicative of the status of the tibial microarchitecture were analyzed.
Materials and Methods
Preparation of the hot water extract of RVS
The hot water extract of RVS was kindly supplied by Lifetree Biotech Co., Ltd (Suwon, Korea) and prepared as described previously [21]. In brief, the timber of RVS was harvested in Wonju, Gangwon-do, South Korea, cut into pieces of 11×1×0.2 cm, mixed with water (1:10, w/v), and then eluted with boiling water (90℃–110℃) for 4 hours. The extract was concentrated to yield 15% solid content. The concentrated extract was diluted with an equal volume of dextrin and dried by a spray-drying method. Urushiol, an allergenic constituent of RVS, was removed and the urushiol-free RVS was authenticated by Korea Advanced Food Research Institute (KAFRI, Seoul, Korea).
Animals
All experiments with rats were acknowledged and regulated by the Institutional Animal Care and Use Committee (IACUC; approved protocol No. P-16-05-A-03) of Konyang University (Daejeon, Korea). Eight-week-old female SD rats (n=48; body weight, 200–250 g) were purchased from Samtako (Osan, Korea). The rats were acclimatized in an environmentally controlled room (temperature, 23±2℃; relative humidity, 60±10%; 12-hour light/dark cycle) and given free access to water and food. All experiments were conducted in accordance with the “Guide for the Care and Use of Laboratory Animals” (National Institutes of Health publication, 8th edition, 2011).
Experimental design and ovariectomy
The rats were randomly divided into the following groups (n=12 per group): sham-surgery (Sham); ovariectomized (OVX); ovariectomized and subsequently administered RVS at a low dose (20 mg/kg; RVS-L); and ovariectomized and subsequently administered RVS at a high dose (100 mg/kg; RVS-H). For the OVX, RVS-L, and RVS-H groups, a ketamine/Rompun cocktail (40 mg ketamine and 10 mg Rompun/kg body weight) was intraperitoneally administered for anesthesia and bilateral oophorectomies were conducted. For the RVS-L and RVS-H groups, RVS diluted in distilled water was intragastrically administered daily for 8 weeks from the first day after the operation. The Sham group was subjected to laparotomy, but without ovariectomy. After the RVS administration period, all rats were weighed and sacrificed. The livers and uterine tubes were excised and weighed. Arterial blood was collected from the abdominal aorta for serological assay. After the separation of the surrounding soft tissues, both tibiae were excised from individual rats and immersed in 10% neutral buffered formalin (NBF) for 24 hours.
Serological assay
After the collection of arterial blood, sera were obtained by centrifugation. The sera were stored at −70℃ until assayed. The levels of serum glutamic-oxaloacetic transaminase (GOT), glutamic pyruvic transaminase (GPT), and total cholesterol were measured with a DRI-CHEM 3000 colorimetric analyzer (Fujifilm, Tokyo, Japan). The serum levels of osteoprotegerin (OPG) and receptor activator of nuclear factor-κB ligand (RANKL) were determined by enzyme-linked immunosorbent assay assay kits (R&D Systems, Minneapolis, MN, USA) using a microplate reader (BioTek Instruments, Winooski, VT, USA) in accordance with the manufacturer's instructions.
Bone µCT
The µCT determination of the excised tibia was performed with a Sky Scan 1076 CT scanner (Aartselaar, Belgium), as previously described [22]. Tibiae fixed with 10% NBF were scanned in batches of three at a nominal resolution (pixels) of 18 µm. The reconstruction was generated by using a modified Feldkamp algorithm using the Skyscan NRecon software. The X-ray source settings were as follows: 70 kV, 100 mA, and a pixel size of 18 µm. A hundred projections were acquired over an angular range of 180°. The image slices were reconstructed using cone-beam reconstruction software (version 2.6) in accordance with the Feldkamp algorithm. The following parameters were analyzed: bone volume per tissue volume (BV/TV), trabecular thickness (Tb.Th), number (Tb.N), and separation (Tb.Sp), and structural indices including trabecular pattern factor (Tb.Pf) and structure model index (SMI).
Hematoxylin and eosin staining and immunohistochemistry
The uterine tubes and tibiae were dehydrated with an ethanol gradient series, embedded in paraffin, and sliced into 5 µm sections using a microtome (RM2255, Leica, Nussloch, Germany). Prior to the tissue processing, the tibiae were decalcified by soaking in 5 ml 10% EDTA at room temperature for 3 weeks; the EDTA solution was changed every 3 days. Decalcification was confirmed by the absence of resistance when the bones were punctured with a pin. Hematoxylin and eosin (H&E) staining of the uterine tubes and tibiae was performed as described in previous studies [2324]. In particular, for preparation of the high-magnification image of tibiae epiphyseal plate, stain with only hematoxylin was used for better visibility. For immunohistochemistry (IHC), the tibiae sections were deparaffinized in xylene, hydrated by immersion in a decreasing ethanol gradient series, and washed in distilled water. Next, each section was incubated with primary antibodies that were diluted in blocking solution at a ratio of 1:200 for rabbit anti-tartrate-resistant acid phosphatase (TRAP; Abcam, Cambridge, MA, USA) for 24 hours at 4℃. After three washes in phosphate-buffered saline, the sections incubated with anti-TRAP were incubated with secondary antibodies containing biotinylated anti-rabbit IgG (Vector Laboratories, Inc., Burlingame, CA, USA) for 1 hour at room temperature. The slides were further incubated with an avidin-biotin complex solution (Vector Laboratories, Inc.) for 1 hour at room temperature. The immunopositive regions turned dark brown after the addition of the chromogen 3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich, St. Louis, MO, USA). Two microscopic fields of tissue sections were randomly selected from each rat (n=12 in each group). These fields were photographed at 200× magnification by a digital camera connected to a light microscope (DM4, Leica) and examined by three blind observers.
Statistical analysis
All data were presented as the mean±standard error of the mean (SEM). Comparisons of the data from the different groups were performed with one-way analysis of variance (ANOVA, PASW Statistics version 18, SPSS Inc., Chicago, IL, USA). The differences with values of P<0.05 were considered to be statistically significant. Each “n” value refers to the number of separate experiments conducted.
Results
RVS treatment did not affect body weight and liver functions, but hyperlipidemia was attenuated in ovariectomized rats
Before the evaluation of the preventive effect of RVS in the rat model of PMO, general conditions and liver functions were assessed in terms of body weight, liver weight, and serological parameters, such as serum GOT and GPT levels (Table 1). Consistent with previous studies [2526], the OVX group exhibited significant weight gain compared with the Sham group. Although there were clear trends of weight reduction in the RVS-L and RVS-H groups compared with that in the OVX group, the differences were not statistically significant. Liver weight, as well as serum GOT and GPT levels, were not significantly increased by ovariectomy or RVS treatment, which suggested that both OVX and RVS did not induce hepatotoxicities such as hepatomegaly. Conversely, total cholesterol, a reliable marker of hyperlipidemia that is commonly associated with postmenopausal syndrome, was significantly elevated in the OVX group; however, treatment with RVS at 20 mg/kg and 100 mg/kg significantly reversed this elevation. Together, these results revealed that the 8-week oral administration of RVS exerted a beneficial effect against hyperlipidemia without causing significant hepatotoxicity.
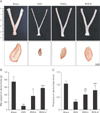
RVS conferred protection against atrophic changes in the uterine tube of ovariectomized rats
The in vivo estrogenic activity of RVS was evaluated by the measurement of the extent of uterine tube atrophies in ovariectomized rats with or without RVS treatment. Representative macroscopic and microscopic images of the uterine tubes of rats in each group are shown in Fig. 1A. The average wet weight and the wall thickness of the uterine tubes of each group are shown in Fig. 1B and C, respectively. As shown in Fig. 1A, compared with the Sham group, the OVX group exhibited macroscopic narrowing of the uterine tubes, which were markedly normalized in the RVS-L and RVS-H groups. As demonstrated in Fig. 1B and C, the RVS-L and RVS-H groups showed a significant reversal of the uterine tube atrophies observed in the OVX group (0.36±0.14 g and 0.57±0.02 g vs. 0.09±0.01 g, respectively, for wet weight; 0.59±0.13 mm and 0.77±0.16 mm vs. 0.31±0.02 mm, respectively, for wall thickness). In particular, the extent of normalization in wet weight and wall thickness of uterine tubes was more prominent in the RVS-H groups than in the RVS-L group, which suggested that the observed changes were dose-dependent.
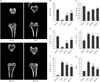
RVS treatment attenuated the deterioration of bone microstructure in ovariectomized rats
The protective effects of RVS on bone microstructure in the rat model of PMO were evaluated by µCT analysis of the excised tibia proximal ends. Representative µCT images of tibial metaphysis confirmed the expected trabecular bone loss in the OVX group compared with the Sham group, whereas marked preservation of the trabecular bone was observed in the RVS-L and RVS-H groups (Fig. 2A). The quantitative analysis of the trabecular responses (Fig. 2B–G) showed that, compared with that in the Sham group, BV/TV, Tb.Th, and Tb.N decreased and Tb.Sp, Tb.Pf, and SMI increased in the OVX group, as expected. RVS treatment of ovariectomized rats dose-dependently increased BV/TV, Tb.Th, and Tb.N relative to that in the OVX group, although all parameters were still lower than in the Sham group. In contrast, Tb.Sp, Tb.Pf, and SMI decreased in a dose-dependent manner in the RVS group in comparison with that in the OVX group. In conjunction with the µCT results on proximal ends of the tibiae, we concluded that RVS exerts a bone-preserving effect against PMO-induced bone loss.
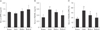
RVS treatment diminished the serum markers of osteoclastogenic activity in ovariectomized rats
As the biology of osteoblasts and osteoclasts is largely regulated by signaling mediators i.e., RANKL and its inhibitor OPG [2728], the serum levels of OPG and RANKL, as well as their relative ratio, were compared at 8 weeks after PMO induction in rats. As shown in Fig. 3A and B, the serum OPG level was not significantly changed in any group, whereas serum RANKL level was elevated in the OVX and RVS-L groups compared with that of the Sham group (0.58±0.07 and 0.50±0.04 vs. 0.33±0.08 ng/ml, respectively). Conversely, serum RANKL level decreased in the RVS-H group. However, the ratio of serum RANKL to OPG decreased in both the RVS-L and RVS-H groups in comparison with that of the OVX group (37.56±4.87% and 29.67±10.41% vs. 51.38±9.76%, respectively) (Fig. 3C). The decrease in serum RANKL to OPG ratio was dose-dependent. Collectively, these results suggested that the protective effect of RVS on bone microstructures was mediated, at least in part, by the suppression of osteoclastic activity.
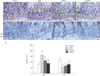
RVS treatment restored osteogenesis and suppressed bone resorption in the tibial epiphyseal plate of ovariectomized rats
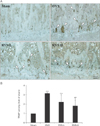
To gain a deeper insight into the underlying bone-preservation effect of RVS, we assessed the possible effects of RVS on osteogenic and osteoclastic activity in the tibial epiphyseal plate, which is actively regenerated via bone remodeling. The H&E-stained sagittal sections (×50) (Fig. 4A) of the proximal tibia showed that the bony trabecular meshwork was apparently reduced and frequently interrupted (as indicated with yellow arrowheads) in the diaphysis in the OVX, RVS-L, and RVS-H groups in comparison with that in the Sham group. As shown in Fig. 4B (×200, representative images depicted with magnified regions of corresponding rectangular boxes in Fig. 4A) and Fig. 4C, the quantitative graph, the animals in the OVX group showed a widened resting zone (RZ) and reciprocally shortened proliferative zone (PZ) in the tibial epiphyseal plate. In contrast, a significant reversal of the effects on the above parameters was observed in the RVS-L and RVS-H groups in comparison with the OVX group (68±8.36 µm and 54±11.40 µm vs. 98±19.23 µm, respectively, for RZ, 40±7.07 µm and 48±8.36 µm vs. 34±8.94 µm, respectively, for PZ). Additionally, the possible effect of RVS on osteoclastic activity was evaluated by IHC using antibody against TRAP, a reliable histologic marker of multi-nucleated osteoclasts. As shown in Fig. 5A (the representative images) and Fig. 5B (the quantitative graph), the TRAP IHC staining (brown stain) showed a robust increase in osteoclast activities (as indicated with white arrowheads), especially in the junction between the diaphysis and the epiphyseal plate in proximal tibia of animals in the OVX group in comparison with that in the Sham group. However, osteoclast activity in the RVS-H group, but not in the RVS-L group, was markedly diminished compared with that of the OVX group (1.81±0.60 vs. 3.16±0.07). Collectively, these findings demonstrated that RVS consumption could reverse or delay the progression of PMO, at least partially, by the simultaneous promotion of osteogenesis and the suppression of osteoclast-mediated bone resorption in the epiphyseal plate.
Discussion
This is the first study to investigate the possible preventive or delaying effects of RVS on PMO progression using various in vivo experiments in a rat model of PMO established by ovariectomy. This study revealed that orally administered RVS (20 or 100 mg/kg for 8 weeks) markedly attenuated the atrophy of the uterine tube, elevation of serum markers of osteoclastic activity (RANKL to OPG ratio), bone microstructural deterioration of the tibia, and imbalance between osteogenic and osteoclastic activity in the tibial epiphyseal plate of the PMO model. Collectively, these results suggested the potential role of RVS as a natural alternative for PMO management.
In addition to the discovery of anti-osteoporotic effects, this study also revealed a serum cholesterol-lowering effect of RVS. In agreement with previous studies, ovariectomy significantly increased serum total cholesterol [293031]. Although the exact mechanisms of ovariectomy-induced hypercholesterolemia have not been clearly elucidated, the most plausible explanation for this phenomenon is the disturbance of cholesterol clearance [32]. Indeed, the ovaries in rodent models are known to be second to the liver with regard to capacity for cholesterol clearance [33]. Post-menopause in elderly women, the incidence of atherosclerosis in coronary arteries surges dramatically owing to E2 deficiency [34]. Increased serum cholesterol level has been implicated as a major predisposition factor for atherosclerosis [35]. Other pharmacologic medications, e.g., statins, which are used to lower cholesterol are often associated with contraindications or side effects, which prevents their use in individuals who are taking other forms of medication. Thus, if possible, alternatives to pharmacologic medication for the reduction of cholesterol level, as well as the risk of atherosclerosis, are highly desirable. The serum cholesterol-lowering effect of RVS demonstrated in this study suggests that RVS may be a safe and efficient alternative to synthetic drugs for the prevention of postmenopausal atherosclerosis.
OPG is an endogenous protein of the tumor necrosis factor receptor superfamily; released by osteoblasts, it inhibits osteoclast activation by acting as a decoy receptor for RANKL [36]. Osteoclasts are multinucleated giant cells derived from hematopoietic progenitors of the monocyte/macrophage lineage through a differentiation process mainly governed by two key cytokines: M-CSF and RANKL [37]. M-CSF induces the proliferation and stimulation of the expression of RANK, the receptor of RANKL, in the precursor cells [38]. The binding of RANK to RANKL leads to the subsequent activation of downstream signaling pathways such as nuclear factor κB (NF-κB) [39]. NF-κB then induces the expression of the transcription factor nuclear factor of activated T cells 1 (NFATc1), a master regulator of osteoclast differentiation [40]. NFATc1 regulates the transcription of several target genes, including TRAP, which participate in the osteoclast phenotype and bone matrix degradation [41]. Thus, the osteoclastic activity might depend predominantly on the expression or activity of RANKL and its inhibitor, OPG, in the bone microenvironment [4243]. The present study showed that RVS decreased serum RANKL, but not the OPG level. Therefore, we can suggest that RVS acts primarily on osteoclasts, but does not directly stimulate osteoblasts to produce OPG, although a detailed study of the mechanism of RVS-mediated RANKL is required.
Phytoestrogens, including RVS, have been known to have multiple modes of action, which indicates that there may be other mechanisms responsible for its anti-osteoporotic effects in addition to the E2-mimetic property demonstrated in the present study; previous studies have demonstrated that various polyphenolic constituents of RVS are potent scavengers of reactive oxygen species (ROS) [4445]. Indeed, although estrogen deficiency has been considered the central and primary cause of osteoporosis in post-menopausal women, a variety of recent studies have indicated that aging and the associated increase in ROS are the main culprits [46]. In addition to original research, one review paper (written in 2010) elegantly provided the paradigm shift from the “estrogen-centric” account of involutional osteoporosis pathogenesis to one in which ROS-related mechanisms intrinsic to bone are the key events [47]. Therefore, although the present study revealed the E2-mimetic property of RVS was at least partly responsible for the protective effects of PMO, as demonstrated by the attenuation of uterine tube atrophy, further investigations will be required to elucidate the exact mechanism from the multiple modes of actions, including the role as a ROS scavenger.

 XML Download
XML Download