

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Despite the economic value of dromedary camels, literature about their reproductive biology is remarkably scarce. Several studies have investigated the histology and histochemistry of the epididymal duct in different mammalian species [1-9]. However, data about the dromedary camel are still relatively limited [10-14].
Camels are seasonal breeders; however, information about their breeding season is rather contradictory [15]. As atypical seasonal breeders, camels exhibit spermatogenesis throughout the year but with a slight reduction compared to that during the breeding season. Consequently, camels may maintain their reproductive capacity throughout the year [16]. Furthermore, both morphometric and histological characteristics of the camel epididymis show slight seasonal differences during rutting and non-rutting seasons [12, 13].
The epididymal epithelium in most mammalian species consists of two main cell types, principal (PCs) and basal cells (BCs), and two accessory cell types, apical cells (ACs) and intraepithelial leukocytes (IELs). However, dark cells (DCs) constitute a fifth cell type reported in the camel epididymis [12-14]. The diverse cellular populations of the epididymal epithelium may account for its wide range of functional capacity. This allows the epididymis to create regionalized and complicated sequential changes in the composition of the luminal fluid throughout its length. This helps the transformation of immature testicular sperm into mature sperm. The maturation process includes morphological and biochemical changes in the sperm plasma membrane in response to the proteins secreted by the epididymal epithelium. Some of these proteins are responsible for inducing progressive motility and for acquiring fertilizing capacity [17-21].
Histochemistry is a biological approach to study the molecular characterization of tissues in relation to their structural organization in situ [22]. In a continuing series of studies on the epididymal duct in different mammalian species, the current work used immunohistochemistry to underscore the spatial distribution of some biologically active proteins within the different regions of the epididymal duct and to highlight the potential structural-functional relationships. A comparative interpretation with other mammalian species (Table 1) [2, 7, 9] was also considered.
The proteins studied were carefully chosen according to their functional relevance; mainly absorption, secretion and contractility. They included angiotensin converting enzyme (ACE), S100, galactosyltransferase (GalTase), alpha smooth muscle actin (α-SMA), connexin-43 (Cx43) and vascular endothelial growth factor (VEGF). ACE is a membranebound glycoprotein, which is detectable in all tissues and body fluids of mammals [23]. It converts angiotensin I (locally produced by epididymal epithelial cells) into angiotensin II, which plays a role regulating electrolyte and fluid transport in the epididymis [24]. S100 belongs to a multifunctional subfamily of Ca2+-binding proteins that have many functions including motility, chemotaxis, and secretion [25]. GalTase is a member of a functional family of enzymes involved in the biosynthesis of glycoconjugate carbohydrate moieties [26]. α-SMA is a contractile protein mainly found in cells with contractile function and is a valuable marker for studying differentiation of smooth muscle cells (SMCs) under normal and pathological conditions [27]. Connexins comprise a large family of trans-membrane proteins that permit intercellular communication [28]. VEGF is a heparin-binding growth factor specific to vascular endothelial cells with potent angiogenic capacity, which is involved in both physiological and pathological conditions [29]. This may be credited to its ability to increase microvascular permeability [30].
Materials and Methods
Animals and tissues
Epididymal tissue specimens were obtained from seven adult clinically healthy, dromedary camels (Camelus dromedarius) slaughtered at the central abattoir in Cairo, Egypt. The epididymal duct was divided into three main parts: the caput, corpus, and cauda epididymis. Specimens were taken from each part of the epididymal duct immediately after slaughter.
Chemicals and methods
Specimens were fixed in Bouin's solution and in a mixture of methanol/glacial acetic acid (2 : 1). Bouin's-fixed specimens were used for routine histological and immunohistochemical staining (ACE, S100, α-SMA, and Cx43). Some proteins (GalTase and VEGF) could not be resolved in Bouin's-fixed sections, and these specimens were fixed in the methanol/glacial acetic acid mixture. Tissue specimens were dehydrated in a graded series of ethanol, cleared in xylene, embedded in Paraplast wax (Sigma-Aldrich, St. Louis, MO, USA) and sectioned at 5 µm thickness. Tissue sections were mounted on positively charged and coated slides (Thermo Scientific, Menzel-Gläser GmbH, Braunschweig, Germany).
Conventional histological techniques
Several conventional stains were used according to standard histological protocols [31] to investigate general histological structure. These included hematoxylin and eosin, Masson's and Goldner's trichrome stains, and the periodic acid-Schiff (PAS) reaction after McManus.
Immunohistochemistry
Dewaxed and rehydrated sections were subjected to inactivation of endogenous peroxidases by an incubation in 1% H2O2 for 15 minutes. Then, the sections were placed in 0.01 M citrate buffer (pH 6) and heated in a microwave oven (700 watts) for 10 minutes for antigen retrieval. The sections were blocked in phosphate buffered saline (PBS) containing 5% bovine serum albumin for 1 hour, and then each section was incubated with its corresponding primary antibody (types, sources, antibody dilutions, and the duration of incubation are shown in Table 2) in a humidified chamber. The sections were washed three times in PBS for 5 minutes and incubated with biotinylated secondary antibodies (types, sources, and dilutions are shown in Table 2) for 30 minutes at room temperature. The sections were washed in PBS for 10 minutes. Then, the secondary antibody was detected using the Vectastain ABC kit (Vector Laboratories Inc., Burlingame, CA, USA). First, each section was covered with a 100× dilution of A and B reagent in PBS (1 µl reagent A, 1 µl reagent B, and 98 µl PBS), washed three times in PBS for 10 minutes, and color was developed using DAB reagent (Sigma-Aldrich). Sections were counterstained with hematoxylin for 30 seconds, washed in water, dehydrated through graded ethanol, cleared in xylene, and mounted with DPX permanent mounting media (Sigma-Aldrich).
Positive and negative controls
Immunohistochemical negative controls, in which the primary or secondary antisera or the ABC reagent was omitted, produced no positive staining. Positive controls were used according to the instructions provided by the manufacturers of the primary antibodies.
Labeling assessment and photomicrography
A semi-quantitative subjective scoring was used by three independent observers to assess the immunolabeling. Photomicrographs were taken using an imaging system consisting of a light microscope (Leica DM LB, Leica Microsystems, Wetzlar, Germany) and a digital camera (Leica EC3, Leica Microsystems Ltd., Heerbrugg, Switzerland).
Results
Histological findings
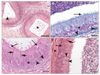
A cross section of the camel epididymal duct at the caput region revealed an irregular contour that varied from triangular to stellate-shaped lumina containing no or few spermatozoa (Fig. 1A). In contrast, the lumina at the corpus and cauda were more regular and were generally oval or circular. Unlike the case in the caput region, the lumina possessed many spermatozoa (Fig. 1B, D). All three epididymal duct regions were lined by pseudostratified columnar epithelium. PCs and BCs were seen along the entire length of the duct (Fig. 1). The apical borders of the PCs had stereocilia, which exhibited a weak to moderate PAS reaction (Fig. 1C). In addition to these cell types, ACs were variably seen in different regions of the duct (Fig. 1C, D). DCs appeared among the PCs as narrow, tall, and darkly stained cells, extending from the basement membrane to the lumen (Fig. 1A). DCs had dark, elongated, and fusiform nuclei and were observed in all epididymal segments but their frequency increased toward the cauda. Additionally, IELs were found throughout the entire length of the epididymal duct. The epithelium contained intraepithelial glands, whose lumina were surrounded by simple columnar or cuboidal cells. Although intraepithelial glands were observed mainly in the distal part of the corpus, they were infrequently seen in other parts.
The epididymal epithelium was surrounded by thin lamina propria (Fig. 1B) and a peritubular muscle coat (PMC) of numerous layers of circularly and obliquely oriented SMCs. The interstitium contained loose connective tissue in which the epididymal blood and lymph vessels and nerve fibers were distributed. Many different cell types were found in the interstitium. These cells were mainly fibroblasts as well as macrophages, lymphocytes, and plasma cells.
Immunohistochemical findings
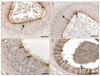
ACE: Apical surfaces of most PCs in several tubules at the caput of the epididymis displayed intense ACE-immunoreactivity (IR) (Table 3, Fig. 2A, B); however, some tubules expressed partial staining or were completely negative. Both ACs and DCs expressed variable IR ranging from negative to strongly positive (Fig. 2A, B). BCs were mainly negative. The epithelium lining the corpus region displayed variable IR ranging from negative to strongly positive. With the exception of ACs, which expressed strong ACE-IR, all other cell types showed variable IR ranging from negative to strong binding (Fig. 2C). Most tubules at the cauda epididymis showed moderate to strong reacting PCs alternating with totally negative PCs (Fig. 2D). Stereocilia expressed ACE-IR weakly. DCs showed variable staining ranging from negative to moderate IR. BCs, IELs, and stereocilia of the PCs failed to express any IR throughout the length of the duct.
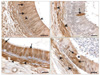
S100: Some tubules at the caput region displayed moderate S100-IR. The epithelium showed moderate and intense binding in PCs (cytoplasm and nuclei) and DCs, respectively. The PMC was moderately reactive (Fig. 3A). Other tubules exhibited somewhat weaker cytoplasmic IR, but the nuclei displayed variable reactivity ranging from negative to strongly intense. BC nuclei expressed negative to weak IR. Stereocilia of PCs, IELs, and ACs were mostly negative. At the corpus region, all other parameters were absolutely negative except for strong S100-IR expressed by PMC and some ACs (Fig. 3B). Some tubules showed intensely positive PCs alternating with negatively reactive ones at the cauda (Fig. 3C). Although PCs displayed negative reactivity in other tubules, many other strongly reactive PCs were found bordering the intraepithelial gland lumina (Fig. 3D). Similarly, the ACs manifested a reaction ranging from negative to moderately intense. Additionally, PMCs displayed consistent IR.
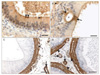
GalTase: The epithelium lining the caput epididymis showed mostly negative BCs and weak to moderate GalTase-immunoreactive PCs. The apical surface of the epithelium presented strongly reactive ACs (Fig. 4A). Although most PCs at the corpus region were negative, some PCs bordering the intraepithelial glands expressed strong IR (Fig. 4B). Reactivity at the cauda was similar to that at the caput but was rather weaker, stereocilia of PCs, BCs, DCs, and IELs were negative along the length of the duct, whereas the vascular endothelium generally showed moderate to strong positive reactivity (Fig. 4A, B).
α-SMA: A consistently strong immunostaining for α-SMA was expressed by the peritubular and the vascular SMCs in all epididymal regions (Fig. 4C, D).
Cx43: No binding sites or IR were found in any epithelial or interstitial structures along the duct.
VEGF: VEGF-IR was entirely absent from both epithelial and interstitial structures along the whole length of the duct.
Discussion
The results of the present study revealed that the epididymal epithelium in the dromedary camel, similar to the other species, consists of the well-known PCs, BCs, ACs, and IELs [2, 6, 32, 33]. Additionally, DCs were also found in the camel epididymal epithelium in agreement with previous work [12-14].
The variable regional S100-IR displayed by PCs agrees with previous studies in the ox [2], donkey [7], and buffalo bull [9] epididymis. Additionally, ACs exhibited obvious S100-IR in the different epididymal regions. Similar findings were reported for buffalo bulls and the donkey, but not in the ox. As a multifunctional subfamily of Ca2+-binding proteins, S100 has a wide range of diverse functions [25]. Although the exact biological role of S100 in the epididymis is unknown, it is thought to be involved in the absorptive and secretory functions of the intra-testicular excurrent duct system [34]. Similarly, S100 may promote comparable tasks in the camel extra-testicular excurrent duct system. In disagreement with the findings reported in the epididymis from ox [2] and European bison [35], no S100-IR was observed in endothelia lining the blood and lymph vessels.
The current findings indicate that the luminal surfaces of the PCs showed remarkable ACE reactivity, particularly at the caput and cauda. ACE binding sites were localized in some tubules and absent in others. This variation may reflect sub-regional functional differences. Similar results were reported in humans [36], ox [2], and donkey [7], but not in buffalo bulls [9]. In contrast, camel epididymis stereocilia displayed a somewhat weaker reaction. Moreover, ACs exhibited a variable reaction ranging from negative to strong ACE binding throughout the whole length of the duct. These findings agree with the results reported in buffalo bulls and donkey, but contradict the case in ox that exhibit no ACE reactivity in ACs. It is evident that ACE converts angiotensin I, locally produced by epididymal epithelium, into angiotensin II. Angiotensin II regulates electrolytes and fluid transport in the epididymis [24]. Furthermore, the vascular endothelium, mainly consisting of subepithelial blood vessels, expressed moderate ACE reactivity. Similar findings were reported in epididymis from humans [36], ox [2], donkey [7], and buffalo bulls [9]. It is worth mentioning that endothelial ACE may participate in regulating vascular tone and, in turn, control blood flow [37, 38] through epididymal tissues.
PCs of the epididymal epithelium at the caput and corpus regions of large ruminants [2, 9] express strong to moderately intense GalTase-immunostaining, particularly in stereocilia and the Golgi zone (GZ). Unlike the case in large ruminants, PCs in camel epididymis exhibited a different distribution pattern represented by a well-distinct reaction in the apical surfaces of only some PCs but not in the GZ or stereocilia. These findings agree with those reported in the donkey [7]. In the present work and in agreement with the findings reported in large ruminants, but not in the donkey, the epithelium lining the cauda was entirely negative. Similarly, ACs expressed moderate to strong GalTase-IR. This disagrees with the case in ruminants but agrees with that in the donkey, confirming distinct species differences. Moreover, the PCs surrounding the intraepithelial glands, mainly at the camel corpus epididymis, exhibited strong GalTase-IR. In contrast to large ruminants [2, 9], it seems that the PCs of the camel caput epididymis are not the main secretory cells, and that the ACs and the PCs bordering the intraepithelial glands might co-play such a role. Although the significance of the epididymal and seminal plasma GalTase activity is unknown, the enzyme may be implicated in glycosylation events that are important during gamete interaction [39]. Notably, GalTase belongs to a functional family of enzymes that are responsible for the biosynthesis of glycoprotein carbohydrate moieties [40]. Alterations in sperm membranes may result from the incorporation of glycoproteins, which are of epididymal origin [18, 20, 21].
In agreement with previous work on the epididymal duct in the ox [2] and buffalo bulls [9], no VEGF-IR was observed in epididymal epithelium of the camel. In contrast, VEGF is expressed in the BCs of donkey epididymis [7], in the BCs and certain peritubular cells in humans [41], and in rat [42] epididymis. This variation may be attributed to species differences. VEGF is an angiogenic protein implicated in physiological and pathological conditions [30], which may be due to its ability to increase microvascular permeability [41]. This might be of importance not only for molecular transport but also for migration of mononuclear cells from blood into interstitium and, consequently, into the epididymal epithelium. The leukocytes that occur both in the interstitium and within the epididymal epithelium may participate in the induction of immune tolerance in the male excurrent duct system [43], preventing the initiation of an immune reaction against sperm.
The cytoplasm of the periductal and vascular SMCs showed distinct α-SMA immunostaining. This agrees with the findings reported in the epididymal duct from different mammalian [2, 7, 9, 44] and avian species [44]. α-SMA is very significant to study SMCs differentiation under normal and pathological conditions [27]. Additionally, the periductal SMCs in camel epididymis displayed a variable S100-IR ranging from moderate (at the caput and corpus) to strong (at the cauda) binding. Similar findings were reported for the epididymis from donkey [7] and rodents [45]. These findings agree with those reported for myoepithelial cells in sweat glands [46], in periacinar myoepithelial cells, in periductal SMCs, in the poll gland of male camels [47], and in the vascular smooth muscle in European bison [35]. As intracellular Ca2+-binding proteins, S100 proteins exert several important functions regulating Ca2+ homeostasis and are the key molecules to transduce Ca2+ signaling by interacting with various kinds of target proteins in SMCs to enhance their contractility [25, 35, 48]. The movement of spermatozoa along the epididymal duct is aided by the contraction of peritubular SMCs [49].
Binding sites for Cx43 were not found either within the epididymal epithelium or in the interstitium. Similar results were reported for the epididymis of the ox [2]. In contrast, the current findings disagree with those reported in rat [50], stallion [4] and buffalo bulls [9], which express a positive reaction in the epithelium. This may be another aspect of species variation in the function of the epididymis.
The diverse cell populations in the epididymal epithelium of camels exhibited variable immunostaining for most proteins under study. Accordingly, certain structural-functional relationships might be proposed for each cell type.
Although a general name is given for the most numerous cell populations in the epididymal epithelium, PCs may perform either absorptive or secretory activities or both [51]. This notion is supported by the current findings, which show remarkable ACE-IR on luminal surfaces of the PCs (at the caput and cauda), variable regional S100-IR displayed by PCs, and distinct GalTase-IR on the apical surfaces of some PCs at the caput and in PCs surrounding the intraepithelial glands of the camel corpus. Furthermore, PCs at the cauda displayed alternating IR for both ACE and S100, suggesting different populations of PCs with variable and probably complementary functional capacities.
BCs comprise the second most frequent cell population in the epididymal epithelium. BCs failed to express immunostaining for any of the proteins under study. Though the func tion of BCs is unknown [52], they are assumed to be reserve cells [53] for epididymal epithelium renewal, but this assumption was disproved in previous work on the equine epididymis [54]. However, BCs express positive reactivity with antibodies that recognize intraepithelial macrophages and their transformation into macrophages has been postulated [55].
ACs are another type of cell encountered in the epididymal epithelium, yet they are relatively less frequent than PCs and BCs. Describing ACs with the term "apical cell" is confusing [56], as it has been used to designate different cell types in the mammalian epididymal duct including clear cells, apical mitochondria-rich cells, and a subgroup of PCs with apically located nuclei [57]. ACs have also been called narrow cells [58] and flask cells [57]. DCs are a population of cells in the epithelium lining the epididymal duct in the camel [12, 13]. DCs appear as narrow, tall, and darkly stained cells with dark, elongated, and fusiform nuclei. These characteristics may coincide with the term "narrow cell" or "flask cell," describing some forms of ACs. Because they share expression of ACE and S100 with ACs, DCs might be special forms or certain stages of ACs. Despite several studies on ACs, the uncertainty of their description has delayed a better understanding of their structural-functional characteristics [56]. Thus, the exact functional significance of ACs is not yet known; however, they may be involved in reabsorption and acidification of epididymal fluid [59]. Moreover, the strong reactivity to S100, ACE, and GalTase expressed by ACs in the different regions of the camel epididymis might point to the significance of this cell type for absorptive and secretory activities.
In conclusion, the spatial distribution of different proteins in camel epididymis showed similarities and differences to other mammalian species. No binding sites could be found for either VEGF (similar to large ruminants) or Cx43 (similar to ox). Distinct binding sites for α-SMA and S100 were consistently evident in the periductal SMCs throughout the whole length of the duct. BCs failed to express any IR with any of the proteins under study, whereas ACs expressed moderate to strong immunostaining for S100, ACE, and GalTase. Consequently, ACs are assumed to perform both absorptive and secretory activities. PCs at the cauda displayed probable complementary and alternating IR for both ACE and S100, suggesting different populations of PCs with variable functional capacities.



 XML Download
XML Download