

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Diabetes mellitus (DM) is among the primary risk factors for periodontitis [1]. DM potentiates the severity of periodontitis and further accelerates alveolar bone destruction. Type 2 DM is the most common form of diabetes, characterized by hyperglycemia, insulin resistance, and relative insulin deficiency. Recent studies have demonstrated that bone loss and osteoporotic fracture risk are increased in DM patients [2]. Several mechanisms have been suggested to explain the increased bone loss and fracture risk in DM patients. Abnormal calcium metabolism occurs in patients with DM. Increased urinary excretion of calcium and decreased intestinal calcium absorption can cause a negative calcium balance with secondary hyperparathyroidism and increased bone remodeling, contributing to a lower bone mineral density in DM patients [3]. Diabetic animals produce sufficient amounts of immature mesenchymal tissue but fail to adequately express Runx2 and Dlx5, the transcription factors for osteoblast differentiation [4]. Advanced glycation end products (AGEs) also have been implicated in the development of DM-related osteopenia. In DM patients, a larger amount of AGEs is accumulated in bone tissue. AGEs inhibit the synthesis of bone matrix proteins such as type I collagen and osteocalcin, and decrease mineralized nodule formation in osteoblasts [567]. AGEs have also been suggested as a biomarker for the increased risk of fractures because AGEs have been shown to be associated with bone rigidity [8]. The receptor for AGEs is expressed in bone cells and its activation stimulates the production of reactive oxygen species (ROS), activating the NF-κB signaling pathway [9]. Intracellular ROS-mediated oxidative stress plays a crucial role in bone health. Increased ROS-mediated oxidative stress in osteoblasts and osteocytes contributes to the cell death of osteocytes and mechanical unloading-induced bone loss [1011]. Patients with type 2 DM have been found to show elevated levels of mitochondrial ROS, which contribute to elevated osteolysis and bone fractures in DM patients [12]. Multiple factors, including a negative calcium balance, microvascular damage, AGEs, and ROS, may contribute to the increased bone loss and fracture risk of DM patients, but further studies are necessary to clarify the mechanisms involved in DM-associated osteopenia and fracture risk.
The canonical Wnt signaling pathway controls cell proliferation and differentiation by regulating the degradation and localization of β-catenin. The binding of canonical Wnt ligands to Frizzled and low-density lipoprotein receptor-related protein (LRP5/6) inactivates β-catenin degradation complexes, resulting in the accumulation and nuclear translocation of β-catenin. In the nucleus, β-catenin induces the transcriptional activation of target genes by forming a complex with lymphoid-enhanced factor/T-cell factor (LEF/TCF). Canonical Wnt signaling promotes the differentiation of mesenchymal progenitor cells into osteoblasts. In osteoblasts, the Wnt pathway induces cell proliferation, differentiation, and matrix mineralization, playing a pivotal role in bone development and formation [13]. Sclerostin, a protein encoded by the Sost gene, acts as an antagonist to Wnt signaling by binding to LRP5/6 co-receptors and inhibiting the binding of Wnt ligands [14]. Sclerostin is mainly produced by osteocytes [15]. Sclerostin-mediated inhibition of Wnt signaling affects osteoblast differentiation and bone formation [16]. Recent reports have demonstrated that removing or blocking the function of sclerostin ameliorated alveolar bone defects in an experimentally induced periodontitis model, suggesting that sclerostin plays an important role in periodontitis-associated bone destruction [171819].
We have previously demonstrated that serum sclerostin levels were higher in mice fed with a high-fat diet than control mice, and that tumor necrosis factor α (TNFα) mediates the induction of sclerostin expression in mice fed with a high-fat diet [20]. High fat diet-fed mice were shown to exhibit higher serum glucose levels and glucose intolerance compared to their healthy littermates. Similarly, serum sclerostin levels have been found to be significantly higher in both type 1 and type 2 DM patients independently of gender and age, compared to non-diabetic controls [212223]. These reports suggest that increased sclerostin expression contributes to DM-associated osteopenia and accelerated alveolar bone loss in diabetic periodontitis patients. However, the molecular mechanisms involved in the DM-mediated induction of sclerostin expression remain elusive. In this study, we investigated how hyperglycemia regulates the expression of sclerostin in osteoblasts and osteocytes using cells from the murine cell lines C2C12 and MLO-Y4.
MATERIALS AND METHODS
Materials
The easy-BLUE™ and StarTaq™ reagents were ordered from iNtRON Biotechnology (Seongnam, Korea) and the AccuPower RT-Pre Mix was purchased from Bioneer (Daejeon, Korea). The SYBR Premix ExTaq™ was purchased from TaKaRa (Otsu, Japan). Tissue culture medium and serum were purchased from HyClone (Logan, UT, USA) and BioWhittaker (Walkersville, MD, USA). A Bright-Glo luciferase assay kit was obtained from Promega (Madison, WI, USA). Recombinant bone morphogenetic protein 2 (BMP2) and Wnt3a were purchased from R&D Systems (Minneapolis, MN, USA) and added to the culture medium at a concentration of 50 ng/mL. D-(+)-Glucose and H2O2 were ordered from Sigma (St. Louis, MO, USA).
Cell culture
C2C12 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS), 100 U/mL penicillin and 100 µg/mL streptomycin. The cell culture medium contained 5 mM glucose. In order to induce osteoblastic differentiation, the C2C12 cells were incubated in DMEM supplemented with 50 ng/mL BMP2, 5% FBS, 100 U/mL penicillin, and 100 µg/mL streptomycin after two days of confluence. Osteocytic MLO-Y4 cells were cultured in alpha modified Eagle's medium (αMEM) supplemented with 5% calf serum and 5% FBS.
Reverse transcription-polymerase chain reaction
Quantitative reverse transcription-polymerase chain reaction (RT-PCR) was performed to examine mRNA expression levels. Total RNA was isolated by using an easyBLUE reagent and cDNA was synthesized from the total RNA using the AccuPower RT PreMix, and subsequently used for quantitative real time-PCR amplification using SYBR Premix EX Taq. Each sample was analyzed in triplicate, and the target genes were normalized to the reference housekeeping gene, glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The PCR primer sequences used for real-time PCR were as follows: Sost, (f) 5'-AGCCTTCAGGAATGATGCCAC-3' and (r) 5'-CTTTGGCGTCATAGGGATGGT-3'; TNFα, (f) 5'-GACGACAGCAAGGGACTAGC-3' and (r) 5'-GCTTTCTGTGCTCATGGTGC-3'; Runx2, (f) 5'-TTCTCCAACCCACGAATGCAC-3' and (r) 5'-CAGGTACGTGTGGTAGTGAGT-3'; osterix, (f) 5'-CCCACCCTTCCCTCACTC-3' and (r) 5'-CCTTGTACCACGAGCCAT-3'; alkaline phosphatase, (f) 5'-CCAACTCTTTTGTGCCAG-3' and (r) 5'-GGCTACATTGGTGTTGAGCTTTT-3'; osteocalcin, (f) 5'-CTGACAAAGCCTTCATGT-3' and (r) 5'-GCGCCGGAGTCTGTTCAC-3'; and GAPDH, (f) 5'-TCAATGACAACTTTGTCAAGC-3' and (r) 5'-CCAGGGTTTCTTACTCCTTGG-3'.
Western blot analysis
In order to prepare the whole-cell lysate, cells were lysed in the PRO-PREP (iNtRON Biotechnology) protein extraction solution, which contains protease inhibitors, including PMSF (174 µg/mL), pepstatin A (0.7 µg/mL), leupeptin (2.0 µg/mL), and aprotinin (0.5 µg/mL). The lysates were then sonicated briefly and centrifuged at 16,000 × g for 10 minutes, and the supernatants were used for Western blot analysis. Each sample containing equal amounts of protein was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The proteins separated in the gel were subsequently transferred onto a polyvinylidene fluoride membrane. The membrane was blocked with 5% nonfat dry milk in Tris-buffered saline containing 0.1% Tween20, incubated with sclerostin or actin antibodies, and subsequently incubated with horseradish peroxidase-conjugated secondary antibody. Immune complexes were visualized using the Supex reagent and luminescence was detected with a LAS1000 machine (Fuji PhotoFilm; Tokyo, Japan).
Enzyme-linked immunosorbent assay
Cells were incubated for 48 hours in the medium supplemented with high glucose or H2O2. Whole cell lysates were prepared as described above and the expression levels of TNFα protein in the cell lysates were determined using a commercial enzyme-linked immunosorbent assay (ELISA) kit (Koma Biotech, Seoul, Korea) according to the manufacturer's instructions.
Knockdown of TNFα
siGENOME ON-TARGETplus SMARTpool mouse TNFα siRNA and non-targeting control scrambled siRNA were purchased from Dharmacon (Lafayette, CO, USA). Cells were transfected with siRNA using Dharmafect (Dharmacon, Lafayette, CO, USA) according to the manufacturer's instructions. The efficacy of the knockdown was assessed by quantitative RT-PCR.
Luciferase reporter assays
The transcriptional activity of β-catenin was examined using a Top-Flash luciferase reporter. MLO-Y4 cells were plated into 96-well plates at a density of 2×104 cells/well. The cells were transiently transfected with 0.2 µg of Top-Flash plasmid using the Lipofectamine reagent (Invitrogen, Carlsbad, CA, USA). Cells were then incubated for 72 hours in the presence or absence of high glucose or Wnt3a (50 ng/mL). Luciferase activity was measured using the Bright-Glo luciferase assay kit according to the manufacturer's instructions.
Dichlorofluorescein diacetate assay for measuring intracellular ROS production
Assays were performed as described previously [24], with some modifications. The cells were grown to 100% confluence and incubated in the presence of 2 µM dichlorofluorescein diacetate (DCF-DA) in the dark at room temperature for 10 minutes. The emitted fluorescence was then measured at 490 nm, and this value was used as a baseline for the following experiments. The cells were then incubated in the presence or absence of high glucose, TNFα, or H2O2 for the indicated periods.
Statistical analysis
The data were presented as the mean ± standard deviation. The statistical significance of the results was assessed by one-way or two-way analysis of variation using Prism6 (GraphPad Software Inc., La Jolla, CA, USA). Post-hoc analysis was performed using the Sidak or Tukey's multiple comparisons test to explore the differences among individual means. P-values <0.05 were considered to indicate statistical significance.
RESULTS
High glucose increases sclerostin expression
C2C12 is a mouse myoblast cell line that can be differentiated into osteoblasts by treatment with BMP2 [25]. In order to examine the effect of high glucose on the expression levels of sclerostin, osteoblastic differentiation of C2C12 cells was induced by incubating confluent cells in the presence of 50 ng/mL BMP2. With the initiation of osteogenic differentiation, the C2C12 cells were exposed to varying concentrations of high glucose (40-100 mM) for 48 hours. Quantitative RT-PCR results demonstrated that high glucose increased mRNA levels of sclerostin at all of the concentrations examined, but statistical significance was observed at glucose concentrations higher than 40 mM (Fig. 1A). The fold changes of sclerostin mRNA in the 5, 40, 60, 80, and 100 mM glucose samples were 1.45±0.26, 2.14±0.15, 7.03±1.24, 6.90±0.13, and 15.85±1.71, respectively. However, Western blot analysis showed that the increase in sclerostin protein levels was most significant at the 100 mM concentration (Fig. 1B). Therefore, subsequent experiments were performed using a glucose concentration of 100 mM. High glucose (100 mM)-induced sclerostin expression was also observed in the C2C12 cells after incubation for 24 hours (Fig. 1C).
Since sclerostin is mainly expressed in osteocytes, the effect of high glucose on sclerostin expression was also examined using the MYO-Y4 osteocytic cell line. Similar to the results obtained from the C2C12 cells, 100 mM glucose significantly increased sclerostin expression as assessed by both mRNA and protein levels (Fig. 1D, E). The fold changes of sclerostin mRNA in the 5, 40, and 100 mM glucose samples were 1.00±0.05, 1.38±0.18 and 5.82±0.37, respectively. Interestingly, in MLO-Y4 cells, 40 mM glucose clearly increased sclerostin expression as reflected in the protein levels, but not in the mRNA levels.
High glucose inhibits Wnt/β-catenin signaling
Since high glucose increased the expression levels of sclerostin, we next examined whether high glucose suppresses Wnt/β-catenin signaling. C2C12 cells were incubated for 24 hours in the presence and absence of Wnt3a (50 ng/mL) or high glucose (100 mM), and the expression levels of osteogenic marker genes, including ALP, Runx2, osteocalcin, and osterix, were examined. Wnt3a significantly enhanced the expression levels of all the marker genes examined (Fig. 2A). In addition, high glucose significantly attenuated marker gene expression levels both in basal and in Wnt3a-treated cells (Fig. 2A). The Wnt3a-induced gene expression levels in 5 mM glucose compared to 100 mM glucose were as follows: Runx2, 7.76±1.08 vs. 4.55±0.14; osterix, 29.65±3.87 vs. 16.57±0.82; ALP 21.82±1.76 vs. 9.05±0.14; and osteocalcin, 5.93±0.19 vs. 3.06±0.08.
Since MLO-Y4 cells are osteocytic cells, we directly examined the effect of high glucose on Wnt/β-catenin signaling using the Top-Flash luciferase assay in MLO-Y4 cells, rather than examining the effects on osteogenic marker genes. Since 40 mM glucose increased the sclerostin protein levels in MLO-Y4 cells, we used this concentration to further check the functional effect of high glucose on Wnt/β-catenin signaling. Wnt3a significantly increased Top-Flash activity, which was blocked by treatment for three days with 40 mM glucose (Fig. 2B). The Wnt3a-induced Top-Flash activities in 5 mM glucose compared to 100 mM glucose were 308.50±65.88 vs. 141.50±19.49.
High glucose-induced ROS production contributes to enhanced sclerostin expression
We next examined whether high glucose enhances the production of ROS. Intracellular ROS levels were determined using DCF-DA. H2O2 (1 mM) was used as a positive control. Compared to the control cells, the ROS production was significantly higher in high glucose- and H2O2-treated cells (Fig. 3). The ROS levels in the control, 100 mM glucose, and 1 mM H2O2 groups after two hours of incubation were 141,756±19,110, 257,520±14,679, and 443,844±16,675, respectively. In order to explore whether increased ROS levels contribute to sclerostin expression, C2C12 cells were incubated with 0.1-1 mM H2O2 for 48 hours and the expression levels of sclerostin were determined. Quantitative RT-PCR results demonstrated that H2O2 significantly increased sclerostin mRNA levels in a dose-dependent manner (Fig. 4A). The fold changes of sclerostin mRNA in 0, 0.1, 0.3, and 1.0 mM H2O2 samples were 0.87±0.05, 1.50±0.16, 2.26±0.29, and 3.17±0.30, respectively. In order to further clarify the role of ROS production in high glucose-induced sclerostin expression, C2C12 and MLO-Y4 cells were cultured in the presence or absence of an antioxidant, N-acetylcysteine (20 mM). The addition of N-acetylcysteine significantly suppressed high glucose- and H2O2-induced sclerostin expression as assessed by both mRNA and protein levels (Fig. 4B-E). High glucose-induced sclerostin mRNA levels in the vehicle compared to N-acetylcysteine groups were 3.01±0.20 vs. 1.05±0.10 in C2C12 cells and 2.74±0.35 vs. 0.27±0.04 in MLO-Y4 cells. H2O2-induced sclerostin mRNA levels in the vehicle compared to the N-acetylcysteine groups were 4.18±1.19 vs. 0.41±0.11 in the C2C12 cells and 2.70±0.10 compared to 0.79±0.04 in the MLO-Y4 cells. These results suggest that high glucose-induced ROS production contributes to the enhanced expression of sclerostin.
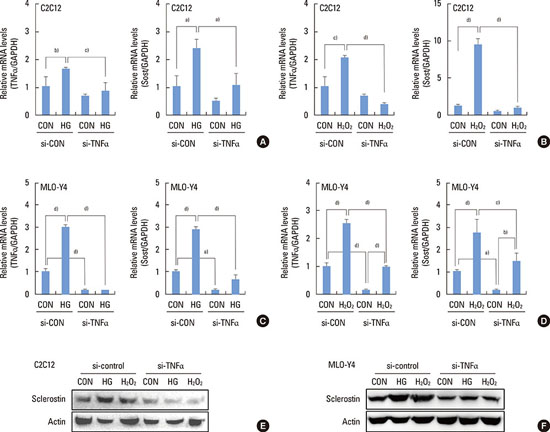
TNFα contributes to high glucose-induced sclerostin expression
Since a previous report has demonstrated that TNFα enhances sclerostin expression in MLO-Y4 cells [20], we examined whether high glucose stimulates TNFα expression. Both high glucose and H2O2 significantly increased the expression levels of the TNFα protein (Fig. 5A, B). Since H2O2 also exerted a stimulatory effect on TNFα expression, we examined whether antioxidant treatment attenuates high glucose-induced TNFα expression. The addition of N-acetylcysteine significantly downregulated high glucose- and H2O2-induced TNFα expression (Fig. 5C, D). High glucose-induced TNFα mRNA levels in the vehicle compared to N-acetylcysteine groups were 18.61±0.62 vs. 7.13±0.68 in the C2C12 cells and 3.25±0.12 vs. 0.32±0.02 in the MLO-Y4 cells. H2O2-induced TNFα mRNA levels in the vehicle compared to N-acetylcysteine groups were 7.74±0.93 vs. 2.54±0.06 in the C2C12 cells and 2.67±0.05 vs. 0.29±0.02 in the MLO-Y4 cells. These results suggest that ROS are involved in high glucose-induced TNFα expression.
In order to further examine whether TNFα mediates high glucose-induced sclerostin expression, the knockdown of TNFα was induced using TNFα siRNA. Quantitative RT-PCR results showed that TNFα siRNA suppressed high glucose- and H2O2-induced TNFα expression (Fig. 6A-D). Compared to control siRNA-transfected cells, high glucose and H2O2-induced sclerostin expression was significantly attenuated in TNFα siRNA-transfected cells (Fig. 6A-D). High glucose-induced sclerostin mRNA levels in the control siRNA compared to TNFα siRNA groups were 2.38±0.35 vs. 1.10±0.41 in the C2C12 cells and 2.90±0.13 vs. 0.64±0.21 in the MLO-Y4 cells. The H2O2-induced sclerostin mRNA levels in the control siRNA compared to TNFα siRNA groups were 9.39±0.92 vs. 1.03±0.07 in the C2C12 cells and 2.75±0.59 vs. 1.46±0.37 in the MLO-Y4 cells. These results suggest that high glucose-induced ROS increase TNFα expression, which subsequently enhances sclerostin expression.
DISCUSSION
Sclerostin is an inhibitor of Wnt/β-catenin signaling and exerts negative effects on bone formation. Hyperglycemia is a representative symptom of DM. Recent studies demonstrated that the risk for osteoporotic fracture and serum sclerostin levels are increased in DM patients and that osteocytic expression of sclerostin is increased in diabetic rats with periodontitis [226]. However, the molecular mechanism of how sclerostin expression is enhanced in diabetic patients remains elusive. In this study, we demonstrated that the exposure of osteoblast lineage cells to high glucose induces ROS and TNFα production, which subsequently upregulates the expression levels of sclerostin.
In this study, we examined the effect of high glucose by adding extra glucose to the culture medium and adjusting the glucose concentration up to 100 mM. In order to rule out the possibility that the high glucose-induced effect on sclerostin resulted from high osmolarity, we compared the effect of high glucose (100 mM) with that of high mannitol (100 mM). Compared to high glucose, the regulatory effect of mannitol on sclerostin and TNFα expression was insignificant, suggesting that the regulatory effect of high glucose on sclerostin and TNFα expression is independent of osmolarity (data not shown). The stimulatory effect of high glucose on sclerostin expression in this study is consistent with the results from a recently published report [27]. In this report, Tanaka et al. [27] demonstrated that treating MLO-Y4-A2 cells with 22 mM glucose for 48 hours significantly increased sclerostin expression, as measured by both mRNA and protein levels.
Chronic hyperglycemia can directly induce an inflammatory state, in which increased levels of cytokines can lead to the degradation of the pancreatic beta cells in both type 1 and type 2 DM [28]. In DM patients, the serum levels of inflammatory cytokines, including IL-6, IL-18, IL-1, and TNFα, are significantly increased [29]. Similar to this in vivo phenomenon, the exposure of C2C12 and MLO-Y4 cells to high glucose significantly increased the expression levels of TNFα mRNA and protein. The induction of TNFα expression by high glucose was blocked by N-acetylcysteine, suggesting that ROS production is necessary for high glucose-induced TNFα expression. TNFα plays an important role in inflammatory bone loss, including rheumatoid arthritis, periodontitis, and osteoporosis [3031]. TNFα also inhibits osteogenic differentiation and bone formation [3233]. Since TNFα also enhances sclerostin expression in a NF-κB-dependent manner in osteocytes [20], it is suggested that enhanced TNFα production by high glucose contributes to increased alveolar bone loss in DM patients.
In DM patients, a greater amount of AGEs is accumulated. The activation of the RAGE receptor by AGEs stimulates the production of ROS and activates the NF-κB signaling pathway [9]. In the present study, high glucose increased ROS production in both C2C12 and MLO-Y4 cells, and the addition of an antioxidant prevented high glucose-induced TNFα and sclerostin expression. In addition, Tanaka et al. [27] demonstrated that AGEs enhance the expression levels of sclerostin in MLO-Y4-A2 cells. Therefore, it is suggested that both AGEs and hyperglycemia contribute to the increase in serum sclerostin levels in DM patients by increasing ROS production.
In conclusion, in this study, it was demonstrated that exposing osteoblast lineage cells to high glucose enhanced ROS and TNFα production, which subsequently induced sclerostin expression. Considering that TNFα and oxidative stress are among the main mediators that enhance osteoclastic bone resorption but suppress osteoblastic bone formation in many inflammatory diseases, the data from in this study further support the hypothesis that the regulation of TNFα and oxidative stress levels in diabetic patients may be of therapeutic relevance for alveolar bone complications derived from chronic hyperglycemia.




 XML Download
XML Download