

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The main function of mitochondria is adenosine triphosphate (ATP) production. Electron transport chains of mitochondria generate H+ electrochemical gradient as electrons pass through complex I, III and IV, activating ATP synthase. The total process is called as oxidative phosphorylation. However, since the 20th century, many studies have focused on the regulatory function of mitochondria in determining the fate of cellular survival and death. Especially, mitochondrial calcium plays a key role in the regulatory mechanism of cell survival and death. Overload of mitochondria calcium during brain insults results in oxidative stress, impaired mitochondrial function, neuronal cell death and neuroinflammation, implicated in pathogenesis of many neurodegenerative diseases [12].
Gunter and Pfeiffer [3] described driving forces of Ca2+ into mitochondria by an equation: ΔµCa=RT ln ([Ca2+]out/[Ca2+]in)+2F(Ψout-#x03A8;in). According to this equation, there are two different driving forces, i) Ca2+ concentration gradient between cytosol calcium ([Ca2+]c) and mitochondrial calcium([Ca2+]m), ii) mitochondria membrane potential (ΔΨm) gradient [3]. Based on correlation between [Ca2+]m and ΔΨm, it has been proposed that pharmacological manipulation of ΔΨm can be a key strategy to prevent neurotoxic mitochondrial overload and neuronal cell death by reducing driving forces of Ca2+ into mitochondria [4]. Regarding these ideas, it has been reported that mild mitochondrial depolarization reduced mitochondrial calcium overload in several studies conducted in different cell types [56].
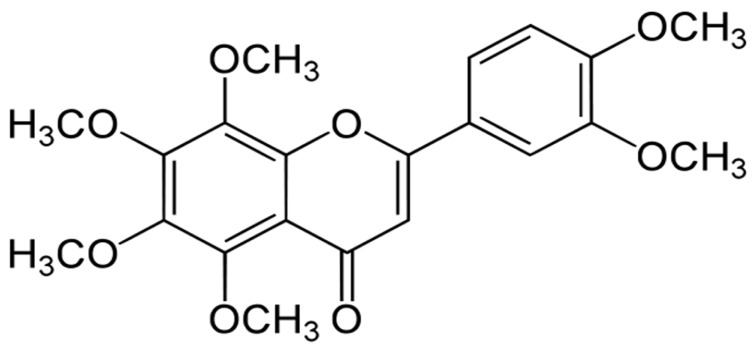
In our previous study, we demonstrated that ethanolic peel extract of Citrus sunki Hort. ex Tanaka (CPE) induces mild mitochondrial depolarization intrinsically [7]. Based on this property, CPE significantly attenuated mitochondrial calcium overload and cell death against H2O2 toxicity in HT-22 cells. Among various flavonoid compounds in CPE (i.e., rutin, hesperidin, sinensetin, tangeretin and nobiletin) belongs to polymethoxylated flavone (PMF) group [7]. Nobiletin exerts several beneficial effects in improving cognitive function or motor deficits in several animal models such as cerebral ischemia [8], Parkinson's and Alzheimer's diseases [910]. In addition, nobiletin induced neurites outgrowth in PC12 cells [11] and significantly suppressed microglial activation and neuroinflammation [12]. These evidences suggest that nobiletin is a promising candidate as a neuroprotective agent and deserves to be thoroughly explored.
Based on these issues, we investigated here whether nobiletin prevents glutamate toxicity-induced neurotoxic mitochondrial calcium overload and neuronal cell death through K+ influx and ΔΨm across mitochondrial inner membrane in primary cerebrocortical neurons or isolated brain mitochondria.
METHODS
Reagents
Tetramethylrhodamine ethyl ester (TMRE), rhod-2 acetoxymethyl ester (Rhod-2 AM), MitoSOX Red, Fura-2 acetoxymethyl ester (Fura-2 AM) were purchased from Invitrogen. Minimal essential medium (MEM), Neurobasal medium, fetal bovine serum (FBS), B-27 serum-free supplement, glutamine and penicillin/streptomycin were purchased from Gibco BRL. Nobiletin was isolated and purified from the peel of Citrus sunki Hort. ex Tanaka in Jeju National University, Department of Biology as described previously [13]. All other reagents were obtained from Sigma-Aldrich, unless otherwise indicated.
Primary culture of cortical neurons
Primary cortical neurons were prepared from cerebral cortices of postnatal 1-day-old Sprague-Dawley rats. The neonatal brain were dissected and the cortices were transferred to plating medium (containing MEM supplemented with 10% FBS, 25 mM glucose, 1 mM sodium pyruvate, 25 mM glutamine and 1% penicillin/streptomycin) and dissociated by trituration using fire-polished glass Pasteur pipettes. Then, cells were plated onto poly-L-lysine-coated round glass coverslips placed in 12-well plates at a density of 1.3×105/well. After 6 h, plating medium was replaced to Neurobasal media supplemented with 2% B-27, 50 mM glutamine and 1% penicillin/streptomycin, and a half of culture medium was replaced every four days. Cultured neurons were incubated at 37℃ in a humidified 5% CO2/ 95% air atmosphere. The cells were used after 7 days in vitro (DIV). The study was approved by Animal Care and Use Committee of Jeju National University and we performed all experiments in accordance with the guidelines.
Dual real-time imaging-based fluorometry of both cytosolic and mitochondrial calcium levels in the same cortical neurons
Fura-2 and Rhod-2 were used to measure cytosolic and mitochondrial calcium levels. Cell-permeable acetoxymethyl ester (AM) forms were used for these two probes. Fura-2 is a ratiometric fluorescent indicator to measure [Ca2+]c using the ratio of emitted fluorescence intensity at excitation wavelengths of 340 and 380 nm [14]. Another calcium indicator Rhod-2, used for a selective [Ca2+]m probe, exhibits charge-driven uptake into the mitochondria and evokes increase of fluorescence upon Ca2+ binding [7]. The cortical neurons cultured on a coverslip were loaded with 10 µM Fura-2 AM and 0.1% Pluronic F127 for 45 min at 37℃. And then, cells were loaded with 2 µM Rhod-2 AM for 30 min at 4℃. After washing three times, a coverslip was transferred to the recording chamber. Cells were continuously superfused with normal Tyrode solution (in mM): NaCl 145, KCl 5, CaCl2 2, MgCl2 1.3, HEPES 10, glucose 10, pH 7.4 with NaOH. The fluorescence was measured with alternative excitation wavelengths of 340 and 380 nm and an emission wavelength of 510 nm for Fura-2, and an excitation wavelength of 540 nm and an emission wave-length of 605 nm for Rhod-2. High-speed filter switching device (Sutter Instrument, Lambda, DG-4) was used for dual recording of Fura-2 and Rhod-2. The fluorescence images were acquired at 6 s intervals using an inverted microscope Olympus IX71 (Olympus) and a cooled-charged device (CCD) camera (Cascade, Roper Scientific). The data were analyzed using Metafluor software (Molecular Devices).
Real-time imaging-based fluorometry of ΔΨm and mitochondrial ROS in intact cortical neurons
The cationic fluorescent probe TMRE is sequestered by mitochondria in proportion to ΔΨm [15]. The cationic MitoSOX Red is also selectively targeted to the mitochondria. Once MitoSOX Red is oxidized by superoxide anions which are the predominant ROS in mitochondria, it exhibits red fluorescence. The cortical neurons were loaded for 15 min at 37℃ with 25 nM TMRE and 5 µM MitoSOX Red for ΔΨm and mitochondrial superoxide, respectively. After then, the neurons on the cover glass were washed three times with normal Tyrode solution. The cover glass was transferred to the recording chamber which was continuously superfused with normal Tyrode solution. Digitized fluorescence images were acquired at 30 s intervals using an inverted microscope Olympus IX71 (Olympus) with a cooled-charged device (CCD) camera (Cascade, Roper Scientific), and analyzed in a personal computer using Metafluor software (Molecular Devices).
Preparation of pure mitochondria isolated from rat brain cortices
The isolated mitochondria were obtained from rat brain cortices, as previously described [16]. In short, the cortices were removed from the brains of 9–16 day-old rats. The fragmented cortices were placed in EGTA-containing isolation buffer (IB) and homogenized in the Dounce-type tissue grinders (Kimble chase). Pestle A and B are used sequentially. The clearance of pestle A and B are 0.0035–0.0065 mm and 0.0010-0.0030 mm respectively. The IB contained (in mM): Mannitol 225, Sucrose 75, HEPES 5, ECTA 3, BSA 0.1%, titrated with KOH to pH 7.4. The homogenates were centrifuged at 600×g for 10 min. The supernatant was transferred to a new tube and then centrifuged again at 600×g for 10 min. Then, supernatants were centrifuged at 12,000×g for 10 min. The pellets were resuspended in IB buffer without EGTA and homogenized using Dounce-type tissue grinders. The clearance of pestle A and B used in this stage were 0.0028–0.0047 mm and 0.0008–0.0022 mm respectively. The homogenates were centrifuged at 12,000×g for 10 min. All the above procedures were carried out at 4℃. The isolated mitochondrial proteins were quantified using Bio-Rad protein assay dye. Electron microscopy (EM) and oxygen consumption rate (OCR) measurement were conducted to evaluate morphology and metabolic activity of isolated mitochondria.
Measurement of ΔΨm and ROS in pure isolated mitochondria using a microplate reader
After preparation of pure mitochondria isolated from rat brain cortices, an isolated brain mitochondrial model was set up. The mitochondria suspension (500 µg of protein/ml) was incubated in recording buffer for 10 min at 37℃ with 25 nM TMRE and 50 µM DCF-DA for ΔΨm and mitochondrial ROS, respectively [71517]. The recording buffer contained (in mM): KCl 100, HEPES 20, Tris 20, NaCl 10, succinate 5, KH2PO4 1, EGTA 0.02, rotenone 0.002, oligomycin 0.001 and CaCl2 0.0001. The loaded mitochondria in suspension were treated with nobiletin or other reagents in 96 well plates. And then, fluorescence intensities were measured using a fluorescence microplate reader (SPECTRA FluoR, Tecan).
Measurement of oxygen consumption rate (OCR)
The OCR of isolated mitochondria was measured using a Seahorse XF-24 extracellular flux analyzer (Seahorse Bioscience) following the manufacturer's protocol. Briefly, 5 µg of isolated mitochondria were suspended in 50 µL of assay medium and transferred to each well for OCR measurement. Mitochondrial assay medium contained (in mM): mannitol 220, sucrose 70, KH2PO4 10, MgCl2 5, HEPES 2, EGTA 1 and fatty acid-free BSA 0.2% (w/v), pH 7.2. One day before the experiment, sensor cartridge was placed into calibration buffer and incubated overnight in a non-CO2 condition at 37℃. The reagents listed below were added sequentially according to the manufacturer's protocol: ADP (2 mM), oligomycin (2 µg/ml) as an inhibitor of mitochondrial ATP synthase, FCCP (2 µM) as an electron transport chain accelerator, rotenone/antimycin A (0.5 µM) as a complex I and III inhibitor. The OCR was recorded by sensor cartridge and analyzed using Seahorse XF-24 software.
Electron microscopy
Electron microscopy for morphological analysis of mitochondria isolated from rat brain cortices was performed as described previously [18]. In short, mitochondria samples obtained through serial centrifugation were fixed overnight with 2.5% glutaraldehyde in 0.15 M cacodylated buffer (pH 7.3) at 4℃, after than samples were centrifuged for 60 s at 3,000 rpm. After washing with the same buffer, samples were post-fixed with 2% osmium tetroxide for 30 min and en bloc stained with 0.1% uranyl acetate solution for 1 h, embedded in 3% agarose gel. Embedded samples were dehydrated through an ascending ethanol series and embedded with an Epon mixture. Thin sections of 70 nm were collected on 200-mesh cooper grid and stained with ueanyl acetate followed by lead citrate. Electron microscopic observation was performed with a Hitachi H-7500 (Hitachi, Japan) transmission electron microscope with 80 kV acceleration voltages.
DPPH free radical scavenging assay
DPPH (1,1-diphenyl-2-picrylhydrazyl) was used to investigate free radical scavenging activity of nobiletin in the solution state, as previously described [19]. The mixture of DPPH and nobiletin was incubated in dark at room temperature for 1 h, and the absorbance was read at 517 nm using a microplate reader (Tecan, Sunrise, AT, USA).
Cell viability assay
MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] was used to investigate cytoprotective effect of nobiletin on cell viability, as previously described [19]. Absorbance was subsequently read at 540 nm using a microplate reader (Model 550, Bio-Rad, USA).
RESULTS
Nobiletin, the key compound of CPE, exhibits neuroprotective effects based on mild mitochondrial depolarization
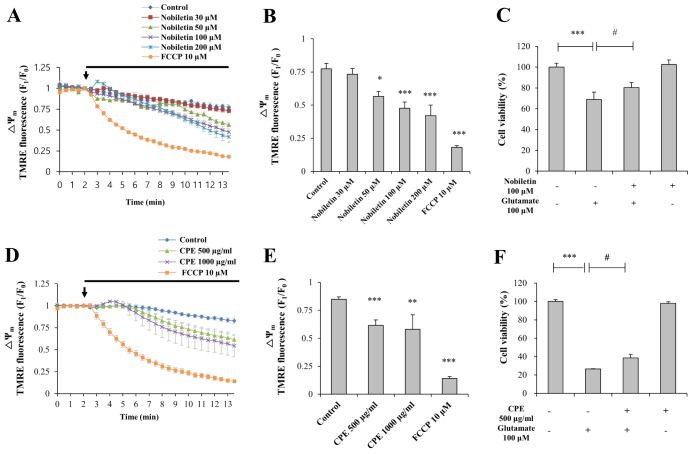
We previously reported that CPE evokes partial mitochondrial depolarization intrinsically while carbonyl cyanide m-chlorophenylhydrazone (CCCP, 10 µM), a well-known ΔΨm dissipation-inducer, evokes complete mitochondrial depolarization [7]. Based on this study, we investigated here whether nobiletin, an active compound of CPE, induces partial depolarization of ΔΨm in resting state and reveal neuroprotective activity against glutamate toxicity in primary cortical neurons (Fig. 2).
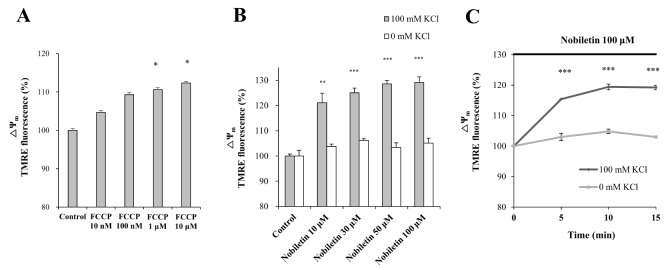
Various concentrations of nobiletin (30, 50, 100 and 200 µM) were superfused over TMRE-loaded cortical neurons on a cover slip in a recording chamber. ΔΨm values were recorded and analyzed using real-time imaging-based fluorometry (see ‘METHODS’ for more detailed description). TMRE fluorescence values from individual cells were normalized to values before drug treatment in Fig. 2A. Recording traces reveal average recordings of TMRE intensities obtained from individual cells. Results demonstrated that nobiletin treatment significantly evoked mild mitochondrial depolarization in dose-dependent manner. Normalized TMRE values at 13-min points were 94.71±5.47%, 73.06±4.79%, 61.47±6.06% and 54.26±10.37% in nobiletin (30, 50, 100 and 200 mM)-treated groups, respectively (Fig. 2B), compared to the control group. Carbonyl cyanide p-(trifluoromethoxy) phenylhydrazone (FCCP) is another ΔΨm dissipation-inducer, similar to CCCP. As shown in Fig. 2, FCCP (10 mM) induced a marked decrease of TMRM values, which means total ΔΨm dissipation. Nobiletin-induced partial mitochondrial depolarization in intact neurons was also confirmed in isolated brain mitochondria using fluorescence microplate reader, as shown in Figs. 5B and C.
The neuroprotective effect of nobiletin against glutamate toxicity was examined in primary cortical neurons using MTT assay. It is well established that glutamate toxicity results in massive and global calcium influx into the cytosol and subsequent mitochondrial calcium overload and evokes cell death [2]. Nobiletin (100 µM) was pretreated for 10 min and glutamate (100 µM) was stimulated for 20 min in the presence of nobiletin. As shown in Fig. 2C, nobiletin significantly increased neuronal cell viability to 80.32±4.80% against glutamate toxicity (100 µM, 20 min).
In addition, we found that CPE (500 and 1,000 µg/ml) intrinsically evoked mitochondrial depolarization in a dose-dependent manner (Figs. 2D and E) and significantly enhanced cell viability (Fig. 2F) against glutamate toxicity (100 µM, 20 min), similar to nobiletin. Results suggest that beneficial effects of CPE on ΔΨm and neuronal cell viability are partially, at least, attributed to its key compound nobiletin, as suggested previously [7].
Nobiletin reduces glutamate-induced mitochondrial calcium overload
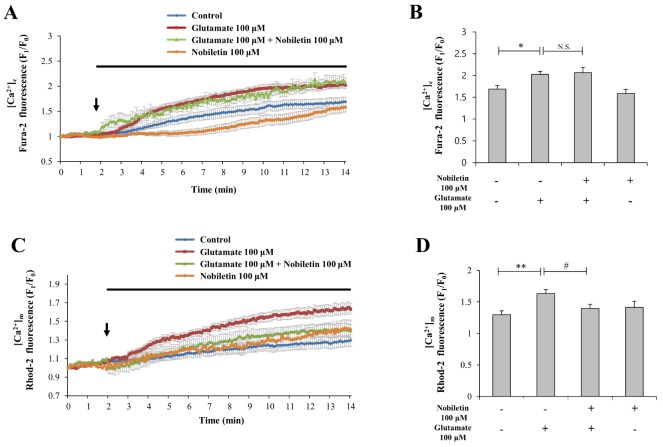
High concentration of glutamate induces excess Ca2+ influx from extracellular space to cytosol via glutamate receptor/channels and subsequently excess cytosol Ca2+ is moved into mitochondria [2]. Mild mitochondrial depolarization attenuates mitochondrial calcium overload by reducing driving force of Ca2+ uptake into mitochondria [456]. Therefore, we investigated here whether nobiletin attenuates mitochondrial calcium overload against glutamate toxicity.
Cytosol and mitochondrial Ca2+ levels were simultaneously recorded and analyzed using dual real-time imaging-based fluorometry with Fura-2 AM and Rhod-2 AM (see ‘METHODS’ for more detailed description). Glutamate (100 µM) in the presence or absence of nobiletin (100 µM) was superfused over Fura-2 AM and Rhod-2 AM-loaded cortical neurons on a cover slip in a recording chamber. Cytosol and mitochondrial Ca2+ levels from individual cells, indicated as Fura-2 AM and Rhod-2 AM fluorescence values, were normalized to values before drug treatment. In Fig. 3A and C, cytosolic and mitochondrial Ca2+ level rises spontaneously in control without glutamate treatment in cortical neuros exposed to external environment in recording chamber. Regarding causes of this phenomenon, several explanations are possible (i.e., decrease of temperature or O2 level, physical stress due to flow system through tubing, fluorometry-related chemical and light toxicities).
Nobiletin markedly abolished glutamate-induced mitochondrial calcium overload in cortical neurons by 85.56±4.10% (Figs. 3C and D). However, nobiletin did not affect glutamate-induced [Ca2+]c increase (Figs. 3A and B). Taken together from Fig. 3, we demonstrated that nobiletin capable of evoking mild mitochondrial depolarization, potently blocked glutamate-induced mitochondrial calcium overload in primary cortical neurons.
Nobiletin attenuates glutamate-induced mitochondrial ROS generation
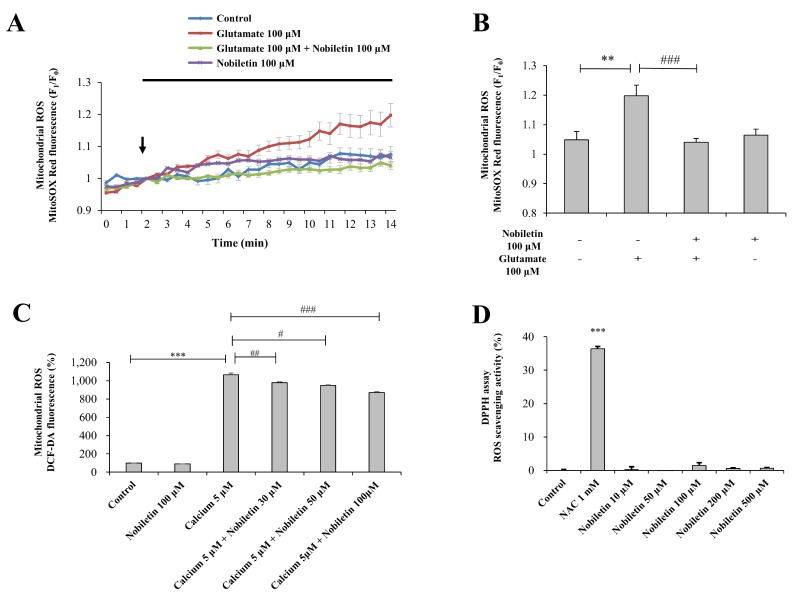
Mitochondrial Ca2+ overload can lead to ROS generation and oxidative stress although Ca2+ has no direct effect in electron transport chains or oxidation/reduction in mitochondria [20]. In this regard, we investigated whether nobiletin inhibits glutamate toxicity-induced mitochondrial ROS generation in intact primary cortical neurons (Figs. 4A and B). In addition, this issue was explored in a pure isolated mitochondria model (Fig. 4C).
In intact cortical neuros, mitochondrial superoxide levels were recorded and analyzed using real-time imaging-based fluorometry with MitoSOX Red. Glutamate (100 µM) in the presence or absence of nobiletin (100 µM) was superfused over MitoSOX Red-loaded cortical neurons on a cover slip in a recording chamber. Mitochondrial ROS levels from individual cells were normalized to values before drug treatment. Nobiletin almost blocked glutamate-induced mitochondrial ROS in intact cortical neurons (Figs. 4A and B).
In the isolated mitochondrial model (see ‘METHODS’ for more detailed description), experiments were conducted with metabolically active and functioning mitochondria as revealed in OCR data (Supplementary Fig. 1B). The cristae structure is well maintained in isolated mitochondria, as revealed in EM results (Supplementary Fig. 1A). High concentration of CaCl2 (5 µM) was treated to isolated mitochondria to mimic glutamate toxicity model in intact cortical neurons. Glutamate treatment does not work in isolated mitochondria since glutamate receptors are expressed in plasma membrane, not in mitochondria membrane. In isolated mitochondrial model as revealed in Fig. 4C, high concentration of Ca2+ was added in the medium instead of glutamate. Using a fluorescence microplate reader, mitochondrial ROS was measured using DCF-DA. Nobiletin significantly attenuated ROS generation in isolated pure mitochondria exposed to high concentration of Ca2+ in a dose (30, 50 and 100 mM)-dependent manner (Fig. 4C), consistent to ROS data from intact cortical neurons.
Data from DPPH free radical scavenging assay revealed that nobiletin did not exhibit direct free radical-scavenging activity in cell-free and mitochondria-free tube system. N-acetyl cysteine (NAC), a well-known anti-oxidant, was used as a positive control in DPPH assay (Fig. 4D). Taken together, results suggest that nobiletin markedly reduces mitochondrial ROS production against glutamate toxicity and mitochondrial calcium overload in intact primary cortical neurons (Figs. 4A and B) and pure isolated mitochondria (Fig. 4C) although it does not have free radical scavenging activity (Fig. 4D).
Nobiletin induces partial mitochondrial depolarization through K+ influx across mitochondrial inner membrane
Influx of K+ from cytosol into the mitochondrial matrix induces mild uncoupling, resulting in dissipation of ΔΨm [4]. The K+ ions are dominant cations among intracellular ions in resting neurons. Based on this background, we hypothesized that nobiletin-induced mild mitochondrial depolarization is mainly mediated by influx of K+ into mitochondria. To confirm correlation between mitochondrial K+ influx and nobiletin effect on ΔΨm, a pure isolated mitochondrial model was used. The effect of FCCP on ΔΨm was investigated as positive control using a fluorescence microplate reader in pure isolated mitochondrial system. When depolarization was induced by FCCP, TMRE intensity increased in dose-dependent manner (Fig. 5A). KCl (100 mM) was replaced with CsCl (100 mM) to remove K+ effects in the medium and thereby K+ dependence in nobiletin effect on ΔΨm was investigated. Figs. 5B and C demonstrated that nobiletin effects on mild mitochondrial depolarization were not shown in absence of K+ in the medium when KCl (100 mM) was replaced to CsCl (100 mM) in an isolated mitochondria model. Results suggest that nobiletin induces partial membrane depolarization by promoting influx of K+ across mitochondrial inner membrane. It could be suggested that K+ influx into mitochondrial matrix by nobiletin is probably mediated by mitochondrial K+ channels such as mitochondrial large-conductance Ca2+-activated K+ channels (mitoBKCa) and mitochondrial ATP-sensitive K+ channels (mitoKATP). Based on this idea, we further explored molecular targets of nobiletin in mitochondria using pharmacological inhibitors. Some preliminary findings related with mitoBKCa and mitoKATP are shown in ‘Supplementary Fig. 2’. However, further detailed studies should be conducted to determine exact molecular targets of nobiletin in mitochondria.
DISCUSSION
In this study, we demonstrated in primary cultured cortical neurons and pure brain mitochondria isolated from rat cortices that nobiletin prevents neurotoxic mitochondrial calcium overload, excess mitochondrial ROS and neuronal cell death through ΔΨm regulation. Results demonstrated that nobiletin evokes partial mitochondrial depolarization in intact cortical neurons (Figs. 2A and B) and isolated brain mitochondria (Figs. 5B and C). Nobiletin markedly attenuated mitochondrial calcium overload and ROS generation in glutamate (100 µM)-stimulated cortical neurons (Figs. 3 C and D, Fig. 4A) and isolated pure mitochondria exposed to high concentration (5 µM) of Ca2+ (Fig. 4C).
Excess exposure of glutamate evoked marked overload of both mitochondria Ca2+ and ROS. Nobiletin treatment significantly reduced these two reciprocal parameters, as shown in Fig. 3 and 4. The exact mechanism by which mitochondrial Ca2+ stimulates ROS generation inside mitochondria remains elusive. However, several plausible mechanisms have been proposed. Possible mechanisms include mitochondrial Ca2+-induced increase of metabolic rate, mitochondrial Ca2+-stimulated dissociation of cytochrome c and mitochondrial Ca2+-stimulated mitochondrial permeability transition pore opening with cytochrome c release [20]. Mitochondrial ROS can inversely affect Ca2+ dynamics and modulate Ca2+ surge. The reciprocal cross-talk between mitochondrial Ca2+ and ROS may results in a feedforward, self-amplified loop evoking subsequent cellular damage and death [21].
The main findings in this study may be mitochondrial K+ influx by nobiletin. We demonstrated that neuroprotective effect of nobiletin via mild mitochondrial depolarization is largely mediated by influx of K+ into mitochondria (Figs. 5B and C). Nobiletin effects on basal ΔΨm were completely abolished in K+-free medium on pure brain mitochondria isolated from rat cortices. Results suggest that K+ influx into mitochondrial matrix is critically involved in the nobiletin effect on ΔΨm. The mitochondrial K+ influx is probably mediated, at least in part, by activation of mitochondrial K+ channels. However, further detailed studies should be conducted to determine exact molecular targets of nobiletin in mitochondria.
There are several ion channels/transporters and electron transport chains on the inner membrane of mitochondria, which are proposed as possible molecular targets of nobiletin in mitochondria. It does not seem that mitochondrial calcium uniporter (MCU) as the major route for mitochondrial Ca2+ uptake is involved in nobiletin-induced ΔΨm depolarization. If nobiletin might attenuate glutamate-induced mitochondrial Ca2+ overload through inhibition of MCU, it should rather hyperpolarize mitochondrial membrane than depolarize it. Several studies have revealed that K+ channels/transporters are present in mitochondria inner membrane: mitoKATP, mitoBKCa, voltage-gated potassium channel Kv1.3, twin-pore domain TASK-3 potassium channels and K+/H+ exchangers. The mitoBKCa and mitoKATP channels are major mitochondrial K+ channels [22]. We demonstrated using pure brain mitochondria isolated from rat cortices that neuroprotective effect of nobiletin via mild mitochondrial depolarization is largely mediated by influx of K+ into mitochondria (Figs. 5B and C). Based on these results, it could be suggested that nobiletin-induced mitochondrial K+ influx is probably mediated, at least in part, by activation of by mitochondrial K+ channels. Based on this idea, we further explored mitochondrial targets of nobiletin. Some preliminary findings related with mitoBKCa and mitoKATP are shown in ‘Supplementary Fig. 2’. We investigated here whether nobiletin-induced ΔΨm depolarization is blocked by iberiotoxin and 5-hydroxydecanoate (5-HD) as well-known inhibitors of mitoBKCa and mitoKATP channels, respectively. As shown in Supplementary Fig. 2, our preliminary findings indicated that nobiletin-induced ΔΨm depolarization was significantly inhibited in the group treated with iberiotoxin (10 nM). These supplementary results could suggest that nobiletin may induce neuroprotective ΔΨm depolarization by promoting the influx of K+ into mitochondria through activation of mitoBKCa. However, further detailed studies should be conducted to determine exact molecular targets of nobiletin in mitochondria.
Among many flavonoids, naringenin, one of flavanones abundant in genus Citrus such as grapefruit and orange, has been widely studied recently. Activation of mitoBKCa is involved in cardioprotective mechanism of naringenin against myocardial ischemia/reperfusion [23]. Naringenin is proposed as one of BKCa channel openers in vascular smooth muscle cells [24]. An electrophysiological study recently revealed that channel activities of mitoKATP and mitoBKCa were enhanced after treatment of 10 µM naringenin in single channel study using mitoplasts isolated from primary human dermal fibroblast cells [25]. These recent reports focus special attention on intracellular pathways mediated by Citrus flavonoids regarding their beneficial effects on several physiological and pathophysiological conditions.
In conclusion, we reveal here that nobiletin evokes partial mitochondrial depolarization in intact cortical neurons and isolated brain mitochondria, and thereby prevents neurotoxic mitochondrial calcium overload, excess mitochondrial ROS and neuronal cell death. Furthermore, K+ influx into mitochondrial matrix is critically involved in the nobiletin effect on ΔΨm. In addition, it could be suggested that the mitochondrial K+ influx is probably mediated, at least in part, by activation of mitochondrial K+ channels. All these findings reveal a beneficial role of nobiletin-induced partial mitochondrial depolarization in neuroprotection, which is similar to ischemic pre-conditioning (IPC) in endogenous mechanism of cardioprotection. Therefore, pharmacological manipulation of ΔΨm through novel substances (i.e., nobiletin) could be a promising strategy to prevent neuronal cell death against brain insults.
 XML Download
XML Download