

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The endoplasmic reticulum (ER) has an important role in protein synthesis, protein maturation, and calcium homeostasis. Biological changes due to energy deprivation and calcium depletion may cause severe ER stress, resulting in the activation of the unfolded protein response (UPR) [1]. UPR triggers an increase in the expression of chaperones that guide protein folding; however, if such a response fails to restore homeostatic balance, apoptotic signaling may occur [2]. Chronic ER stress has been found to be involved in many diseases including fibrosis, atherosclerosis, diabetes, and neurodegenerative diseases [3,4,5,6,7]. An increase in ER stress has been reported in the adipose tissue and liver of diet-induced obese mice and ob/ob mice [8], and islet cells from mice and humans with type 2 diabetes have shown increased ER stress [9]. Biomarkers of the UPR were found to be increased by palmitate in isolated myocytes from diabetic patients, indicating ER stress in the skeletal muscle of diabetic patients in vivo [6].
Exercise is one of the therapies for the treatment of obesity and type 2 diabetes [10], and the known benefits of regular exercise on skeletal muscles include increased levels of glucose transporters [11], antioxidants [12], and fatty acid oxidation enzymes [13]. Muscle contraction during exercise results in the production of reactive oxygen species (ROS) and a subsequent increase in the expression of antioxidant enzymes [14]. This increase of ROS may cause ER stress, resulting in an increase in the production of cellular antioxidants to protect against cell damage. While the relationship between mitochondrial dysfunction and ER stress has been investigated in skeletal muscle tissue of high-fat fed rats in a recent report [15], there have been few studies investigating ER stress in relation to exercise and mitochondrial activity, despite its potential influence on metabolism throughout the whole body.
Recently, several studies have investigated ER stress occurring in muscle tissue during exercise [16,17]. Activation of UPR was observed in human skeletal muscle after a 200-km race [17], and a 4-week period of exercise training resulted in increased UPR as an adaptation to ER stress [16]. Thus, implying that exercise training may induce an ER-stress adaptation in skeletal muscle, and it reported that muscle contraction is directly involved in the activation of the UPR. In addition, mitochondrial biogenesis was also demonstrated to have a key role in controlling UPR [16]. Skeletal muscles require a large amount of energy; as a result, mitochondria play a more important role in skeletal muscle than in other tissues. Therefore, the relationship between mitochondria and ER stress in skeletal muscle was suggested as an area for further study [18]. In addition, the therapeutic effects of exercise and the biological responses it induces are known to be closely related to the intensity of exercise [19]. Moreover, these biological responses vary according to exercise intensity and therefore induce different degrees of ER stress. Thus, we decided to compare ER stress, apoptosis signaling, and mitochondrial biogenesis in low- and high-intensity exercise-trained rats.
To this end, following exercise training in rats, we examined the changes in the level of immunoglobulin heavy chain-binding protein (BiP) in order to detect any disturbances in ER proteins and in the protein kinase RNA-like ER kinase (PERK)-activating transcription factor (ATF) 3/4-C/EBP-homologous protein (CHOP) pathway to assess adaptations to exercise. In addition, we examined changes in the levels of peroxisome proliferator-activator receptor gamma coactivator-1 alpha (PGC-1α) and uncoupling protein 3 (UCP-3) in order to understand the relationship between adaptations to ER stress and mitochondrial-biogenesis gene expression.
METHODS
Animal care and exercise training
Male Sprague-Dawley rats, each weighing approximately 250 g, were purchased from Samtako (Seoul, Korea). The animals were acclimatized for 1 week prior to beginning the experiment by exposure to 12-hour light/dark cycles. The animals treated in accordance with the principles of the Guide to the Care and Use of Experimental Animals of the Yeungnam University Medical Center. The animals were assigned randomly to one of three experimental groups; control (n=7), low-intensity training (LIT, n=7), or high-intensity training (HIT, n=7). Before beginning the exercise protocols, the rats of both exercise groups were adapted to running on a treadmill for 15 min, at increasing speeds from 0~15 m/min, once per day for 5 days. After this adaptation, the rats ran on treadmills with a 10° incline for a period of 60 min on five days per study week. The treadmill speeds were set at 20 m/min in the LIT group and 34 m/min in the HIT group. This experiment was performed as described previously [20]. The trained rats were sacrificed 3 days after the final exercise session to exclude the possible effects of acute exercise stress in the analysis.
Sample collection and analysis
The rats were anesthetized with intra-peritoneal injections of 20 mg/kg tiletamine and zolazepam (Zoletil®; Virbac, Carros, France), and 10 mg/kg of xylazine hydrochloride (Rompun®, Bayer, Monheim, Germany). Food was removed from the cages before sacrifice. After complete anesthesia, the abdominal cavity was opened rapidly following the median line of the abdomen. Blood was drawn rapidly from the abdominal vena cava into syringes. Blood was centrifuged (1800×g for 10 min, 4℃) and the plasma was maintained at -80℃ for the measurement of triacylglycerol (TAG), total cholesterol (TC), glucose, and insulin levels. The gastrocnemius muscle was collected immediately from each rat, and was frozen and stored at -80℃ until the analytical assay was performed. Plasma TAG, TC, and glucose levels were measured using an enzymatic colorimetric assay kit from Sigma-Aldrich (St. Louis, Missouri, USA). Plasma insulin was measured using an enzyme-linked immunosorbent assay kit from Millipore (Billerica, MA, USA).
RNA extraction and gene expression analysis
Frozen tissue samples were homogenized using an Ultra-Turrax T25 (IKA-Labortechnik, Staufel, Germany), and approximately 100 mg of tissue was used for the isolation of RNA. Total RNA was extracted using the guanidine thiocyanate method, and mRNA was purified using a PureLink RNA Mini Kit (Invitrogen, New York, USA) according to the manufacturer's instructions. Total RNA was reverse transcribed in a final volume of 20 µL using a High-Capacity cDNA Reverse Transcription Kit with random primers (Applied Biosystems, Foster City, California, USA) according to the manufacturer's instructions. Reverse-transcribed samples were stored at -20℃. The RNA level during PCR was measured with a continuous fluorescence detector using the Real-Time PCR 7500 system (MJ Research, Waltham, MA) and Power SYBR Green PCR master mix (Applied Biosystems, Foster City, California, USA) according the manufacturers' instructions. To normalize the amount of total RNA present in each reaction, the GAPDH gene was amplified simultaneously. Primers were designed using Primer Express software (Applied Biosystems) and are shown in Table 1. RT-PCR reactions were performed according to the following cycle profile: 10 min at 95℃, 44 cycles for 30 sec at 94℃, 1 min at 58℃, and 2 min at 68℃.
Western blot analysis
The gastrocnemius muscle was homogenized in lysis buffer containing 50 mM HEPES, 1 mM EDTA, 1 mM EGTA, 150 mM NaCl, 50 mM NaF, 1 mM phenylmethylsulfonyl fluoride, 1 mM benzamide, 1 mM Na3VO4, 1 mM dithiothreitol (DTT), 5 mM MgCl2, 1% NP40, 10% glycerol, aprotinin, leupeptin, and pepstatin A. The homogenate was centrifuged at 20000×g for 15 min at 4℃. The protein concentration of the supernatant was measured using the Bradford assay. SDS-PAGE was used to separate 80 µg of protein that was then blotted on a PVDF membrane. The membrane was blocked with 0.1% Tween in tris-buffered saline (TBST) with 5% skimmed milk for 1 h at room temperature. Primary antibodies (PERK, phosphorylated PERK, BiP, CHOP, and GAPDH [PERK, phosphorylated PERK, BiP, and CHOP from Cell Signaling, Danvers, MA, USA; GAPDH from Santa Cruz Biotechnology, Santa Cruz, CA, USA]) were diluted 1:700 in TBST with 5% bovine serum albumin, and were incubated overnight at 4℃. The membranes were incubated with goat anti-rabbit/mouse IgG HRP antibodies (Bio-Rad, Hercules, CA, USA) for 1 h at room temperature then diluted 1:2000 in TBST with 5% skim milk. The membrane was examined using electrochemiluminescence. In this study, GAPDH was used as a control to ensure equal protein loading on the gel and the blots were quantified using MultiGauge software.
RESULTS
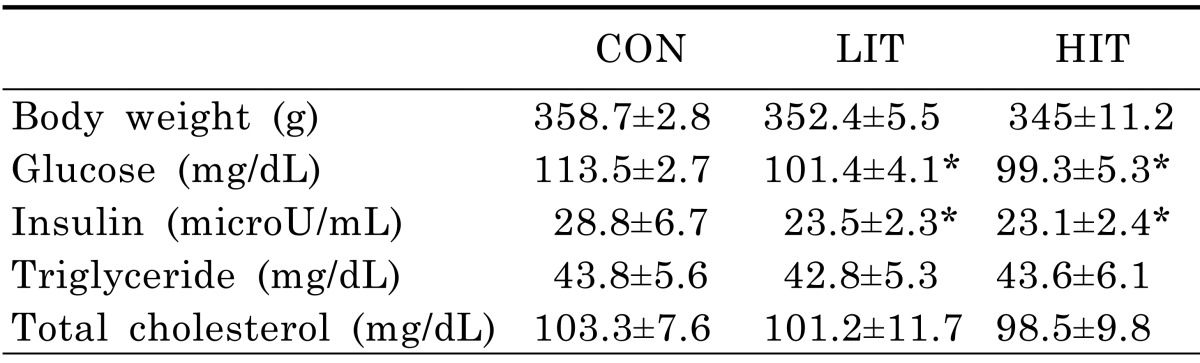
Body weight and blood chemical analysis
Although the average body weights of mice in the LIT (352.4±5.5 g) and HIT (345±11.2 g) groups were lower than in the untrained control group (358.7±2.8 g) after 5 weeks of training, there was no statistically significant difference among the groups (Table 2). The fasting glucose levels of the LIT (101.4±4.1 mg/dL) and HIT (99.3±5.3 mg/dL) groups following 5 weeks were lower than the control group (113.5±2.7 mg/dL). In addition, fasting insulin levels in mice from the LIT (23.5±2.3 µU/mL) and HIT (23.1±2.4 µU/mL) groups were lower than the control group (28.8±6.7 µU/mL). However, there was no significant difference between LIT and HIT groups in terms of fasting glucose and insulin levels. The TAG and TC levels in the fasting state did not show any statistically significant differences across all three groups.
Effects of exercise training on ER stress and UPR in skeletal muscle
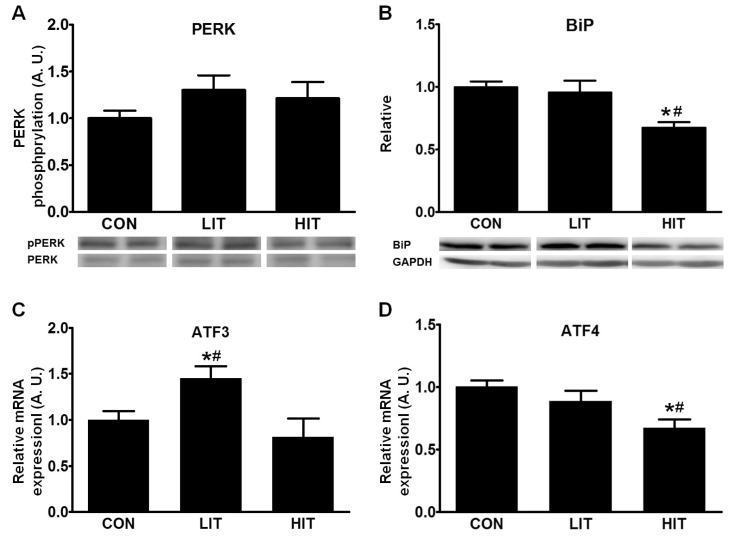
To understand the effects of training intensity on ER stress and UPR in skeletal muscle, we measured PERK phosphorylation, BiP levels and ATF3/4 mRNA expression after 5 weeks of training. Although both intensities of training induced marginal increases in phosphorylation of PERK in skeletal muscle, there were no significant differences across the three groups (Fig. 1A). On the other hand, the HIT group showed significantly reduced BiP levels (32.5%) in skeletal muscle tissue in comparison with the control group (Fig. 1B). LIT-group rats exhibited higher ATF3 gene expression (44.8%) in skeletal muscle tissue than the control group; however, the HIT group did not show any significant difference. Gene expression for ATF4 in the skeletal muscle tissue was significantly decreased in the HIT group (33.0%) in comparison with the control group (Fig. 1C). However, there was no difference in ATF4 gene expression between the control and LIT groups.
Effect of exercise training on the apoptotic pathway in skeletal muscle
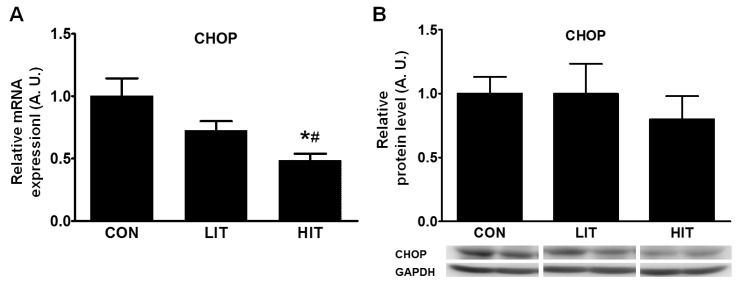
Conditions ranging from unsuccessful UPR to chronic ER stress can induce apoptosis; therefore, changes in the apoptotic pathway were examined using the levels of CHOP protein and CHOP mRNA. We found a 51.4% decrease in CHOP gene expression in the HIT group in comparison with the control group (Fig. 2A). The level of CHOP protein was somewhat decreased in the HIT group but the difference between the groups was not significant (Fig. 2B).
Gene expression of mitochondrial biogenesis in skeletal muscle after exercise training
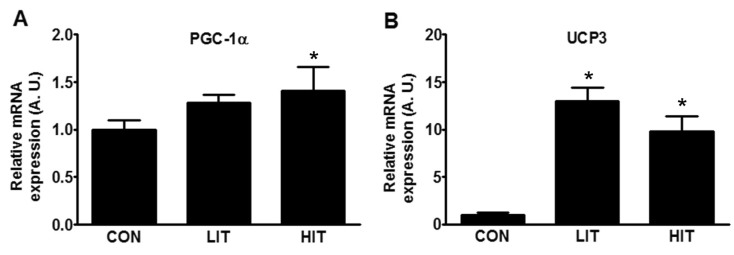
Owing to certain differences in UPR that were dependent on the intensity of exercise training, we attempted to assess the physiological changes in skeletal muscle during training. Mitochondrial biogenesis was measured by the expression levels of the proteins PGC-1α, and UCP3. An increase in the mRNA expression of PGC-1α of 41.5% was found in the HIT group in comparison with the control group (Fig. 3A). The expression of UCP3 in both the trained groups increased; it demonstrated approximately 13-fold (LIT) and 10-fold (HIT) increases in comparison with the control group (Fig. 3B).
DISCUSSION
We observed ER stress, apoptosis signaling, and mitochondrial biogenesis in the skeletal muscle of rats following LIT and HIT to clarify the adaptation mechanisms in muscle physiology induced by training. The duration of training (5 weeks) was selected in order to avoid focusing on the metabolic benefits associated with long-term training, such as with a 12-week training program [20,21]; this study focused on the effects of exercise training on skeletal muscles. The 5-week duration is similar to that of recent studies investigating ER stress and exercise training [16,22]. In previous studies, ER stress markers in the skeletal muscle in ob/ob mice and high-fat fed mice were unchanged by training [8], and 2 weeks of muscle unloading in the skeletal muscles of rats also did not induce UPR [23]. However, more recent studies demonstrated definite UPR in the skeletal muscle of mice [16] and humans [17]. After a 200-km run, levels of BiP, ATF4 and XBP1 mRNA increased in comparison with basal level [17]. These results demonstrated a definite role for the UPR in skeletal muscle physiology during short-term exercise and training.
There are three main effectors of ER stress: ATF6, inositol-requiring enzyme 1 alpha, and PERK [1]. Activated PERK, a type I ER transmembrane protein kinase, inhibits general protein translation in cells through the inactivation of the initiation factor eIF2a [24]; prolonged activation of PERK can result in either cellular adaptation to ER stress or apoptosis [25]. Our results from muscles after 5 weeks of training did not show any statistically significant increase in PERK phosphorylation, suggesting the adaptation of muscle cells to ER stress by repeated exercise; in addition, there was no indication of significant ER stress at the time of sacrifice. Therefore, the observed UPRs can be considered to be the result of adaption to exercise training and not the result of acute exercise. The Hsp70 ER chaperone, BiP, is considered an indicator of disturbances in ER protein homeostasis, and the accumulation of unfolded/misfolded proteins can result in the release of BiP from three effectors in order to transmit signals of ER stress [26,27]. BiP was found to be increased in the skeletal muscles of humans after a long distance run [17] but there were no significant changes in BiP levels in the skeletal muscles of mice following 4 weeks of training [16]. BiP levels in the LIT group in our study did not differ from those in the control group. However, BiP levels in the HIT group were significantly decreased in comparison with the control and LIT groups. This implies that the adaptive response was greater, resulting in decreased ER stress in skeletal muscle, following high-intensity exercise training.
In this study, HIT repressed the expression of ATF4 in comparison with the control and LIT groups. This translates to decreased ER stress in skeletal muscle following 5 weeks of HIT. It is known that during UPR, ATF4 upregulates genes that are involved in amino acid uptake, glutathione biosynthesis, and resistance to oxidative stress [28]. In a previous study, the gene expression of ATF4 was increased after a single exercise session, but ATF4 expression was decreased after 4 weeks of exercise training [16]. We cannot explain the reason for the difference in ATF3 gene expression between the LIT and HIT groups; LIT increased ATF3 in comparison with the control and HIT groups. However, this result implies that the ATF3 and ATF4 may be involved in different pathways leading to adaptation to exercise training.
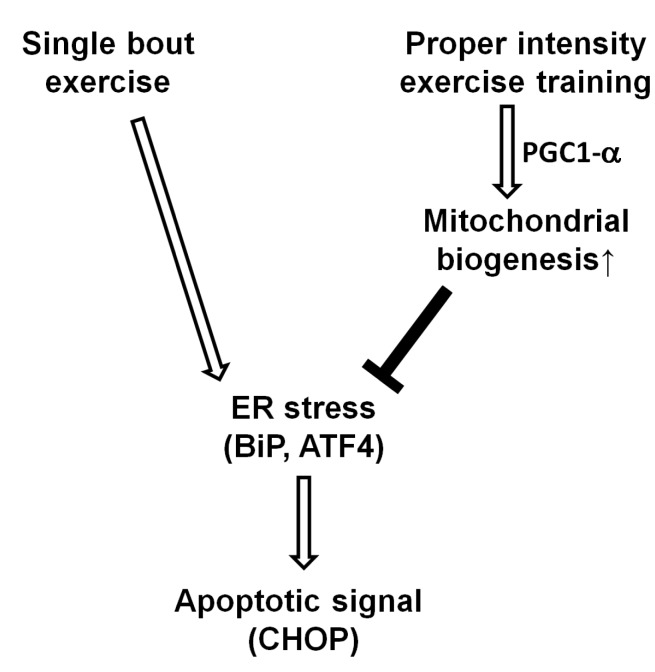
Adaptation to exercise training was also observed in the levels of the protein CHOP. If the UPR to ER stress is unsuccessful, chronic ER stress can induce ER stress-related cell death [29]. Although there are several known pathways of apoptosis resulting from ER stress, CHOP is the bestknown transcriptional factor of ER-induced cell death [26]. In addition, ATF3 and ATF4 are known to activate CHOP [30], which has pro-apoptotic activity through the downregulation of BCL-2 and upregulation of BIM, PUMA, and GADD34 [4]. Intriguingly, the level of CHOP mRNA was significantly reduced in the skeletal muscle of the trained rats and the decrease in gene expression was greater in the HIT group in comparison with the LIT group. However, while the level of CHOP was somewhat decreased in the trained groups, the difference was not significant. These results imply that regular exercise with proper intensity may diminish apoptotic signaling in skeletal muscle, and it can be considered as a protective mechanism for upcoming ER stress (Fig. 4). This protective effect seems likely to be derived from a decrease in ATF4 levels; this is consistent with a recent report that showed that the knock-down of ATF4 decreased CHOP, and subsequent apoptosis [31]. However, we were unable to detect any change in another important transcription factor, spliced XBP1 mRNA, which is also involved in ER stress-induced apoptosis (data not shown).
ER stress is known to induce mitochondrial dysfunction [8,32] and mitochondrial damage, observed by Southern blot analysis and mitochondrial copy number, is known to be associated with ER stress in skeletal muscle [15]. The expression of PGC-1α was increased in the HIT group in comparison with the control group; this is concordant with a recent report that PGC-1α is one of the key factors in the regulation of the UPR observed in mice [16]. Oxidative stress is a well-known factor in inducing ER stress [5] and is considered a local messenger between the ER and mitochondria [32]. However, exercise is known to induce ROS scavengers such as superoxide dismutase and catalase [14]. Although we did not observe the ROS and their scavengers, adapted UPR and increased mitochondrial gene expression may be related to oxidative stress in our study. In addition, ROS have been reported to be involved in the regulation of PGC-1α in skeletal muscle [33] and antioxidant supplementation has been observed inhibiting the exercise-induced increase in PGC-1α [34]. Therefore, we can assume that the ROS generated during exercise training stimulated mitochondrial biogenesis and contributed to the adaptation to, and preconditioning against, further damage. UCP1 and UCP3 are known to respond rapidly to biological stimuli such as energy depletion, and the level of UCP3 in a trained human did not show any increase after acute exercise [35]. Therefore, relatively lower gene expression of UCP3 in the HIT group in comparison with the LIT group seems understandable as a more adapted status is reached in skeletal muscle as a result of more intense training. Misfolded protein can be degraded by the ubiquitin-proteasome pathway and autophagy [5]; as a result, further investigation of these two pathways will provide more information to gain clearer insight into exercise training and the adaptation mechanisms in skeletal muscle. Although the role of PGC-1α in mitochondrial biogenesis is well known, it is also involved in fiber-type switching, enhanced fatty acid oxidation, and angiogenesis [21]. Therefore, the relationship of these effects with ER stress should also be the focus of further study.
In summary, we found that 5-week high-intensity exercise training resulted in increased mitochondrial biogenesis, and decreased ER stress and apoptotic signal in the skeletal muscle tissue of rats (Fig. 4).

 XML Download
XML Download