

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
As a major allergen of the house dust mite Dermatophagoides pteronyssinus (D)., Der p 2 is one of the most important allergenic specificities associated with the cause of allergic asthma.1 It is a highly polymorphic protein with an MW of 15-kDa belonging to the Myeloid Differentiation 2 (MD-2)-related lipid-recognition (ML) family.234 In addition to its strong IgE reactivity, there is evidence that Der p 2 binds to lipopolysaccharide (LPS), a ligand of MD-2, and that Der p 2-LPS induces proinflammatory responses through TLR4,5 similar to the human MD-2-LPS complex.5
More than 13 isoforms of Der p 2, called variants by the IUIS nomenclature, are known to exist with polymorphisms being demonstrated in several different countries.346 There is a dominant polymorphic pattern that occurs from prevalent substitutions at the 4 residues of the canonical Der p 2.0101, namely, V40L, T47S, M111L, and D114N.3 To date, 7 variants with different combinations of these substitutions have been described.3 Of these 7 variants, 4 (Der p 2.0103, 0104, 0109, and 0110) contain 114N and a combination of dominant polymorphic residues at V40L, T47S, and M111L.3 Only Der p 2.0104 has all 4 of the dominant polymorphic residues differing from Der p 2.0101.3 The reactivity of a monoclonal antibody has shown that the substitutions at residue 114 produces an antigenic change;7 however, variants with and without this substitution have been shown to have higher IgE-binding activity than Der p 2.0101.89 These results suggest that further studies on IgE binding to the variants could reveal information useful for developing hypoallergenic Der p 2 and determining if variants should be considered for immunotherapy based on the induction of blocking antibodies.
In addition to structure-function relationships, the geographical distribution of different variants might change the nature of the IgE response in different regions. The Der p 2 variants found in the study area of Australia were different to those found in Thailand,3 and variants with 114D have not yet been found in Bangkok,35 while some were found in Perth where environmental Der p 2.0101 is prevalent.35 The type of IgE binding found for house dust mite (HDM)-allergic individuals in different countries could thus be different and if blocking antibodies are required, this could affect immunotherapy. Since anti-Der p 2 antibodies typically account for 25% of anti-mite IgE antibodies,1 the variant could affect serological diagnostic criteria.
Therefore, this study aimed to examine possible IgE-binding consequences of naturally found variants with substitutions in the 4 dominant polymorphic residues, and determine if IgE-binding affinity to the variants would differ in the study areas of Australia and Thailand where D. pteronyssinus is the dominant cause of sensitization and where a different pattern of amino acid substitution has been described.
MATERIALS AND METHODS
Allergens
Yeast-expressed recombinant Der p 2 (rDer p 2) was produced as previously described10 from a single colony of transformed P. pastoris KM71H strain containing Der p 2 variant cDNA and grown in buffered complex media containing glycerol (BMGY) in a ratio of 1:100 at 30℃ with shaking until O.D.600nm of 20.0-25.0. Cells were harvested and resuspended in a solution containing 1% yeast, 2% casamino acid, 1% asparagine, 0.34% Yeast Nitrogen Base, 4×10-5 % biotin, and 0.5% methanol (YCM) at a 10-fold more concentration and were grown for 72 hours at 30℃. The expression of the rDer p 2 variants was induced by adding 0.5% methanol to media every 24 hours. All secreted rDer p 2 variants were purified through a size-exclusion Superdex 75 column (GE Healthcare Biosciences AB, Uppsala, Sweden), precipitated in trichloroacetic acid, and redissolved in 100 mM NaPO4/150 mM NaCl at pH 7.0, and stored at -20℃ until used. Natural Der p 2 (nDer p 2) was produced from spent mite media as previously described.10
Determination of protein concentration
Protein concentration was determined using the absorbance value at 280 nm and calculated from the following equation.11 This equation assumes that all the cysteines form disulphide bonds, where n is a number of indicated amino acid per polypeptide, and M is molecular weight.
Circular dichroism (CD) spectroscopy
Protein samples were prepared at a final concentration of 10 µM in PBS. CD spectra were recorded from 260 to 190 nm at 0.02-cm path length and 1-nm resolution, with a scan speed of 50 nm/minute on a Jasco J-700 spectropolarimeter. The CD data was used to calculate secondary structures with the CDPro suite of programs.12
Hydrophobic staining of Der p 2 with 1,8-ANS
A protocol to characterize the hydrophobic cavity of rDer p 2 has been described.10 According to this protocol, 1-anilinonaphthalene 8-sulfonic acid (1,8-ANS) (Sigma, St. Louis, MO, USA) binding to measure the hydrophobic binding capacity of Der p 2 was conducted as before10 with 1,8-ANS dissolved in methanol at a concentration determined spectrophotometrically using a molar extinction coefficient of 8,000 M/cm at 372 nm. For assay, 3.5 µM natural Der p 2 (nDer p 2) or rDer p 2 variants were mixed with 200 µM ANS for 10 minutes at 25℃. Assay was performed on a Jasco FP-6300 fluorometer. ANS was excited at 390 nm with 5-nm slit width, and the emission spectra were measured at 400-600 nm with 5-nm slit width.
Sera
The use of sera from skin prick-positive HDM-allergic donors for IgE binding was approved by the Institutional Review Board at Siriraj Hospital (SiEc459/2008), Thailand and the Princess Margaret Hospital Ethics Committee, Perth, Western Australia (1347/EP). The mouse anti-rDer p 2.0101 and anti-Der p 2.0110 were each a pool from 4 primed and boosted highly responding C57BL/6 mice immunized according to the Telethon Kids Animal Ethics Committee (Project 201).
Direct antibody binding
First, 500 ng of each Der p 2 variant in PBS was added to a 96-well Maxisorb plate (Nunc, Rochester, NY, USA) and incubated at 4℃ overnight, and then 1:8-1:32 diluted sera from HDM-allergic donors in PBS were added and incubated at room temperature for 2 hours. Thereafter, 1:1,000 diluted HRP-labeled goat IgG anti-human IgE antibodies (KPL, Gaithersburg, MD, USA) in PBS containing 0.05% Tween 20 was added and incubated at room temperature for 2 hours. Bound antibody was detected with a chromogenic substrate, 3, 3', 5, 5'-tetramethylbenzidine (TMB) (KPL, Gaithersburg, MD, USA), and absorbance was recorded at 450 nm after 10 minutes or for the Perth sera by the Wallac DELFIA® assay using Europium-conjugated streptavidin (PerkinElmer, Waltham, MA, USA) as previously described.8
Inhibition ELISA
As previously described,10 sera were diluted 1:8 to 1:32 and incubated with serially diluted rDer p 2 variants or nDer p 2 in PBS containing 0.05% Tween 20 at 4℃ overnight. The absorbed sera were centrifuged at 17,210 g for 10 minutes before the supernatant was added to a 96-well plate, precoated with Der p 2.0104, and incubated at room temperature for 2 hours. Then, 1:1,000 diluted HRP-labeled goat IgG anti-human IgE antibodies (KPL) in PBS containing 0.5% Tween 20 was added and incubated at room temperature for 1 hour. Bound antibody was detected with a TMB or DELFIA® as mentioned above. The results were calculated as mean and SD of the percent inhibition obtained for each of the different sera, and IC50 was calculated from a regression curve constructed with these values.
Inhibition ELISA of mouse-specific polyclonal IgG antibodies against Der p 2.0101 was performed with the same protocol as above except that a dilution of mouse antibody was based on a predetermined titer by direct binding ELISA and 96-well plate was precoated with Der p 2.0101. Bound mouse IgG was detected by 1:1,000 diluted HRP-labeled goat IgG anti mouse IgG (H+L) antibodies (KPL) in PBS containing 0.5% Tween 20 was added and incubated at room temperature for 1 hour. Bound antibody was detected with a TMB as mentioned above.
RESULTS
Secondary structure and accessible hydrophobicity
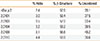
Five rDer p 2 variants with different amino acids in positions 40, 47, 111, and 114, namely, 0101, 0103, 0104, 0109, and 0110 (Table 1) were purified and shown to migrate in SDS-PAGE as discrete 15-kDa bands that, as shown previously,10 bound to monoclonal anti Der p 2 antibodies (data not shown). CD was performed to calculate the secondary structure compositions. All variants had a similar composition of secondary structure, and these were similar to the structure content determined for nDer p 2 (Table 2) noting the expected high percentage of β-sheet.
The accessible hydrophobicity that, from the known structure of Der p 2, could be attributed to their hydrophobic cavities was analyzed from emitted fluorescent intensity of cavity-bound hydrophobic probe ANS. Der p 2.0103 emitted the highest value (-337) of fluorescent intensity at λmax 473 nm, whereas Der p 2. 0101 emitted the lowest value (-97) of fluorescent intensity at λmax 484 nm (Fig. 1). Both the Der p 2.0104 and 0109 emitted fluorescent intensity at the same λmax 473 nm, with values of 222 and 255, respectively (Fig. 1). The nDer p 2 emitted fluorescent intensity (-237) at λmax 481 nm (Fig. 1). The addition of acrylamide to the ANS-stained rDer p 2 variants did not quench the emissions showing that the fluorescence was likely to be from the ANS in the cavity of the molecules (data not shown).
Relative IgE-binding affinities
Binding affinity of IgE antibodies to the Der p 2 variants was measured with panels of individual sera from HDM-allergic subjects in Bangkok, Thailand and then HDM-allergic subjects in Perth, Western Australia. As shown in Table 1, the sequences of Der p 2.0104 were found in Perth and Bangkok, while the sequences of Der p 2.0101 (and similar variants) were found in Perth but not in Bangkok and Der p 2.0109 and 0110 were found only in Bangkok. The ability of the different variants to inhibit the binding of IgE antibody to the Der p 2.0104 (the highest known IgE-binding variant) was determined for each individual in each panel, then the IC50 was calculated for the Bangkok and Perth sera from regression lines. Der p 2.0101 was the worst inhibitor in Bangkok with its IC50 being 3-fold higher than those of Der p 2.0104 and 0109, and 7.5-fold higher than that of Der p 2.0103 (P<0.01 by one-way ANOVA) (Fig. 2A). For the Perth subjects, Der p 2.0101 and Der p 2.0104 had similar IC50 values, while the IC50 for Der p 2.0110 (the only non-Perth variant tested) was 7-fold greater (P<0.05) (Fig. 2B).
Direct IgE binding to variants
The direct IgE antibody titer of the individual sera of HDM-allergic subjects in Bangkok and Perth were determined using assays where the allergen variants were used for coating the microtitre plates. Since the total anti-Der p 2 titer of the individuals varied, the differences that could be attributed to the variants were appraised by calculating the binding to each variant as ratio of the binding to Der p 2.0101, the allergen that is generally used worldwide for rDer p 2 (Fig. 3). The IgE binding of the Perth sera exhibited conspicuous differences between different variants, showing that the binding to Der p 2.0104 was significantly higher (ratio of Der p 2.0101 to Der p 2.0104, 0.46) than that to Der p 2.0101 (P<0.01, one-way ANOVA).The ratios of binding of Der p 2.0103, 0109, and 0110 were over 1 compared to Der p 2.0101, showing the titers were lower than this variant (P<0.001, P<0.001, and P<0.01, respectively). For the IgE binding of the Bangkok sera, there was a similar trend toward higher binding to Der p 2.0104 (P<0.01) and lower binding to Der p 2.0103, 0109, and 0110.
When the binding to the variants was compared between the 2 populations, the ratio of IgE binding to the 0101/0103 was lower in sera from Bangkok than from Perth (P<0.05), and that to the 0101/0109 ratio was also lower in sera from Bangkok (P<0.05) (Fig. 3).
Variant-binding specificities of mouse antisera
The aforementioned results suggest that differences in IgE binding to different variants may not be simply due to differences in exposure to each variant. To study this possibility, the ability of different variants to block the binding of IgG antibody from a pool of sera of mice immunized with Der p 2.0101 to precoated Der p 2.0101 was tested (Fig. 4). The weakest inhibitor was Der p 2.0101 itself with lower IC50 values found for Der p 2.0104 and 0109 and very high inhibition by Der p 2.0103.
However, comparison of IgG binding to the variants directly coated on the plates (Fig. 5A) indicated that more antibodies from the anti-Der p 2.0101 bound to the Der p 2.0101 than to all the other variants with Der p 2.0101 showing less than 30% of the titer. Antiserum reciprocally produced against Der p 2.0110 bound preferentially to itself and Der p 2.0109 (Fig. 5B).
DISCUSSION
The recombinant variants with different combinations of amino acids in the dominant polymorphic pattern of V40L, T47S, M111L, and D114N substitutions had a similar overall content of the secondary structure as determined by CD analysis, which was similar to nDer p 2 with all containing a high percentage of β-strand. However, the accessibility of the hydrophobic cavities of the variants varied as indicated by the different fluorescent intensity emitted by the bound ANS hydrophobic probe. It has been proposed that the formation of the hydrophobic cavity of Der p 2 would be changed upon ligand binding and that its motion would be like a clamshell-like motion.14 Interestingly, ANS-staining of the canonical Der p 2.0101 had the lowest fluorescent intensity indicating a less accessible cavity, while other variants had high fluorescent intensity indicating better accessibility than that found for the natural Der p 2 preparation which is known to at least contain Der p 2.0102 and 0103.5 Substitutions to residues 40, 47, 111, and 114, which are found together on a loop region of Der p 2 and are neither very surface exposed nor part of the cavity, induce structural changes that affect ANS binding and, as discussed below, surface changes detectable by serology. It should be noted that although the results describe variations in IgE reactivity, the IC50 values, even when different, were in the ng/mL range for inhibition experiments, indicating largely intact structures.
Profound differences were found when the affinity of the antibody cross-reactivity was measured by the ability of the variants to inhibit the binding of mouse IgG anti- Der p 2.0101. Der p 2.0103 (LSMN) had the highest affinity (IC50 of -0.07 µg/mL). However, affinities to Der p 2.0110 (VSMN), Der p 2.0104 (LSLN), and Der p 2.0109 (VSLN) were 1-2.6 µg/mL, with the lowest to Der p 2.0101 itself, with an IC50 of -6.9 µg/mL being 10-fold less than that to Der p 2.0103. While it is not possible to ascribe different IC50s to a particular amino acid substitution, the differences in the IC50 between the variants with only N at position 114 clearly show that substitutions other than the 114 exchange detected by monoclonal antibody are important. The finding that inhibition by Der p 2.0101 had the highest IC50 could reflect the fact that homologous inhibition is more difficult to achieve because of different epitopes or high-affinity binding to the solid phase. This was demonstrated by the direct binding experiment in which Der p 2.0101 bound more than the other variants, especially Der p 2.0110 which had only 30% of the titer. The direct binding of antiserum produced against Der p 2.0110 was reciprocal, although not as marked results with the highest binding to the immunogen. Comparison of the IC50 values of Der p 2.0103 and Der p 2.0110 (Fig. 4) which differ only at residue 40, suggests that at least when methionine is present at position 111, the V40L can play an important role in antibody reactivity. The V40L located inside the molecule interacts with several residues.15 However, although valine and leucine are hydrophobic residues, differences in the size of sidechain may alter the structure of a particular epitope. As for the M111L, our results suggested that both residues M111L and D114N may be the components of the epitope that has been demonstrated by other studies.16
The IC50 experiments showed that the variants affected IgE antibody cross-reactivity, whose nature differed between Bangkok and Perth. The inhibition of IgE binding of sera from Bangkok to Der p 2.0104 with rDer p 2.0101 required 90 ng/mL to reach IC50, while a comparable result was achieved at almost 10-fold less concentrations in the experiments with Perth sera where blocking by Der p 2.0101 and Der p 2.0104 was similar. This corresponds well with the result that Der p 2.0101-like variants or any variants with 114D were not found in the study of 17 variants from homes in Bangkok.3 In contrast, such variants were readily found in homes in Perth.5 A similar phenomenon may have been evident in Perth where Der p 2.0110, a variant not found in Perth, had a significantly higher IC50 than Der p 2.0101 and 0104 demonstrated in any environment. The other variants not found in Perth, Der p 2.0103 and Der p 2.0109, showed similar IgE binding ratios compared to Der p 2.0101 in direct binding assays, so further testing to determine if their inhibitions are similar could be instructive. It should be noted that since our experiments only compared the inhibition of binding with high IgE binding to Der p 2.0104, the marked differences do not represent affinities determined by measuring homologous inhibitions. The direct binding analysis gives a measure of the total antibody binding to each variant, including cross-reactivities to other variants. IgE from Bangkok sera less bound to Der p 2.0101 than to Der p 2.0104, and less binding to Der p 2.0109 and 0110 was found in the environment. The direct binding for Perth showed, as reported before,8 that IgE binding to Der p 2.0104 is higher than that to Der p 2.0101, with higher binding in the environment where the Der p 2.0101 can be readily detected. This suggests some intrinsically greater immunogenicity for Der p 2.0104 or greater cross-reactivity to antibodies produced against other variants. The new information in this paper for Perth shows that the IgE-binding titer to Der p 2.0103, 0109, and 0110, which have not been found in this environment, are all lower than those to Der p 2.0101 and Der p 2.0104, and that the differences are more marked in Perth than in Thailand. Another difference between the Perth and Bangkok regions that should be taken into account is that 30% of the acarofauna of Bangkok is Dermatophagoides farinae,16 whereas they are not found in Western Australia.18 Der f 2 has L, T, V, and N at positions equivalent to 40, 47, 111, and 114, and they have not shown polymorphisms.318
Analysis of Der p 2 variants and their effects on IgE binding in this study could not be done without recombinant proteins. With respect to the recombinant proteins, investigations of natural variants are indeed indicated, but it should be noted that recombinant Der p 2 is being used in the studies of human IgE binding. It has been shown that component-resolved diagnosis in HDM allergy can resolve different types of allergic sensitization and disease associations in HDM allergy192021 and answer the question whether subjects with possible cross-reactions are allergic at all.20 Clinical trials with rDer p 2 are also being investigated.22 The different serological responses of subjects from the Perth and Bangkok regions also indicate that natural variants within the environment have shaped the fine specificity of the anti-Der p 2 antibody response, although there are other differences between the regions. With regard to the variant detected, the absence of Der p 2.0101-like variants in the 17 Bangkok sequences3 compared to the 8/9 Der p 2.0101-like variants found in Perth5 is very significant, even though finer analysis of the variants would require more comprehensive studies. Clearly, acknowledgment of the environmental variants found in the highly investigated major population center of temperate regions would be useful, and the results here show the potential for such investigations.
This study showed that the 4 dominant polymorphic residues may be important factors affecting IgE-binding affinity and the degree of binding to Der p 2 variants as well as the specificity of IgG antibodies produced following immunization. Basophil tests have found that natural variants, including the most disparate Der p 2.0101 and Der p 2.0104, induce almost identical degranulation,17 which would be expected from our finding showing that although they were different, the antibody titers to all the variants were high. Our study on IgE binding and competition, however, indicates that targeting the 4 residues with less conservative substitutions could be a strategy for developing hypoallergic Der p 2. The T-cell repertoire to Der p 2 is complex with individuals responding to several epitopes,23 so variants made hypoallergenic by a few amino acid substitution would be expected to retain most of their T-cell reactivity as in folding variants, such as those described for Der p 124 that could lose some epitopes by altering antigen processing. This would need to be tested as per the Der p 1 study.24 Studies on IgE binding and competition also show that it would be important to determine which of the Der p 2 variants would induce IgG antibodies that can block IgE binding to the variants in the environment. This would be critical for immunotherapy based on the strategy for inducing IgG-blocking antibodies. For new types of therapy with recombinant constructs as well as the current therapy with allergen extracts, it would be optimal for a recombinant allergen cocktail or the extracts to contain the same variants as those in the environment. The higher IgE binding to Der p 2.0104 in both Bangkok, Thailand and Perth, Western Australia indicates that Der p 2.0104 would be a better reagent than Der p 2.0101 for IgE binding tests, although for highly allergic subjects, as already shown,817 both would be expected to bind to IgE and mediate mast cell degranulation.
In conclusion, the affinity of IgE antibody cross-reactivity, direct IgE binding, and the specificities of antibodies induced by vaccination show that determination of allergic sensitization to Der p 2 can be optimized with the knowledge that variants and IgE responses differ among regions with different variants. IgG antibodies with blocking activity would be optimally produced with variants known to be important in the environment.






 XML Download
XML Download