

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Atopic dermatitis (AD) is a common chronic inflammatory skin disease affecting 15-25% of children and 3% among adults.1-3 It is strongly associated with asthma and allergic sensitization.1-3 Recent data showed that AD is a major problem in developing, as well as developed countries.1 Approximately 85% of patients with AD begin during early childhood, and 70% of patients severe AD develop asthma or allergic rhinitis later in life.4-6 Patients with severe or persistent AD and their families suffer from significant impairment in their quality of life.7 In addition, AD places a heavy economic burden not only on patients and their families, but also on society in general.8,9 AD skin is characterized by immune dysregulation and epidermal barrier defects such as abnormal terminal differentiation of keratinocytes and decreased cornification.6,10-12 This review will focus on structural abnormalities in the epidermis and their modulation by immune responses.
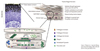
Normal epidermal barrier
The epidermis provides a physical and permeability barrier (Figure).13-15 This barrier is continuously regenerated by terminally differentiating keratinocytes, a process known as cornification or keratinization. Epidermal differentiation begins with the migration of keratinocytes from the basal layer, and ends with the formation of the cornified layer.13,14 Human epidermis undergoes complete turnover every 28 days.16 Cell proliferation, differentiation and death occur sequentially, and each process is characterized by the expression of specific proteins including Filaggrin (FLG), Loricrin (LOR) and Involucrin (IVL) that are cross linked to form a impermeable skin barrier.13
Profilaggrin is a member of the fused S100 family of S100 Ca2+-binding proteins and a large (-500 kDa) complex protein.13,14 It is expressed in the granular layer of epidermis and consists of a unique N-terminal domain; a region with multiple FLG repeats; and a unique C-terminal end domain.13,14 During terminal differentiation at the granular to cornified cell transition, profilaggrin is rapidly dephosphorylated and cleaved by several endoproteases including caspase-14, to generate FLG and the N-terminal domain.13,17,18 FLG aggregates the keratin filaments into tight bundles.13,14 FLG proteolysis occurs upon exposure to a low humidity environment and might be inhibited by high humidity.19 In the cornified cell, FLG is degraded into free amino acids by caspase-14 and these amino acids are essential for the retention of water contributing to the osmolarity in the cornified layer.17,18
LOR is expressed in the granular layer during cornification and an insoluble protein.13,14 LOR is also one of the main components of the epidermal envelope and is intermixed with profilaggrin, comprising 80% of the total protein mass of the cornified layer.20 LOR seems to function as a major reinforcement protein for the cornified envelope.14,21 IVL is a common component of the cornified envelope and consists of repeating peptide units.22 IVL is an early component in the assembly of cornified envelops and provides a scaffold to which other proteins subsequently become crosslinked.14 In the cornified-envelope structure, IVL is adjacent to the cell membrane.14
Roles of epidermal barrier
The epidermis serves as the first line of defense against invading pathogens and allergens.3,23,24 The epidermal cornified layer is 15 to 29 nm thick and composed of structural proteins and a specialized lipid layer.25,26 Various investigators have demonstrated that a defective skin barrier enhances allergen sensitization, leading to systemic allergic responses such as increased IgE levels and airway hyper-reactivity.27,28 FLG deficiency alters the shape of corneocytes in the human skin,12 and exhibits skin inflammation and enhances epicutaneous sensitization in murine models of eczema.15 In addition, it has been shown that FLG deficiency confers a paracellular skin barrier abnormality that reduces inflammatory thresholds to irritants and haptens.12
On the other hand, the absence of LOR in null mice results in no obvious abnormality, although an early, temporary deficiency in water-barrier function is observed.29 This indicates that keratinocytes could compensate for the loss of LOR by using other available proteins to form a functional cornified layer.30 The specialized lipid layer in the conrnified envelope also provide a barrier against water and prevent water loss.11,12,26
FLG mutations and AD
FLG mutations are the best known causes of impaired skin barrier and is considered as predisposing factors for AD.2,31,32 Many genes have been associated with AD and these genes include components of the epidermal skin barrier.33 R510X and 2282del4 are the most common mutant alleles associated with AD in Northern European populations with 10 to 50% of patients with AD having 1 of 2 loss-of-function mutations in FLG.11,34-37 The FLG mutations are currently considered as a major risk factor for AD, particularly in patients who have onset of AD at 2 years or younger and patients with persistent AD.31,38 It has been reported that an individual having one FLG null mutation is at four-fold greater risk of having early-onset AD, while an individual with two mutant FLG allels has an ~80-fold increased risk of developing AD compared with an individual with normal FLG alleles.39 The FLG null alleles identified in asian populations are also significant predisposing factors for AD.40,41
However, it is likely that aside from FLG mutations, there will like be other causes of epidermal barrier defects in AD. In this regard, a significant number of patients with AD do not have any of the known FLG mutations, and conversely, approximately 40% of individuals with FLG-null alleles do not develop AD.37 In addition, up to 50% of patients with AD have FLG mutations, and those who have the mutation eventually recover from the disease.42 Therefore, more work is needed to identify other causes of epidermal abnormalities in AD beyond FLG mutations, and why some individuals never get AD despite the fact that they carry these mutations.
Immune dysregulation of epidermal barrier in AD
The pathophysiology of AD is not well understood, although it is clear that gene-environment interactions in genetically predisposed individuals play a central role. Investigators have shown a highly significant association between abnormalities in the epidermal barrier and risk of developing early-onset, severe and persistent AD.31,35,43 Of note, these may be due to both mutations of genes encoding proteins such as FLG, as well as modulation of epidermal protein levels by Th2 cytokines.44,45 The Th2 cytokines interleukin (IL)-4 and IL-13, which are overexpressed in the acute skin lesions of AD patients, can down-regulate FLG, LOR and IVL, potentially further exacerbating both the epidermal skin barrier defects.44,45 A critical link between the barrier defect in AD patients with FLG mutations and Th2 polarization could be explained in part by enhanced allergen penetration through the damaged epidermis accompanied by increased production of thymic stromal lymphopoietin (TSLP) by keratinocytes leading to a Th2-type milieu.46 Importantly, patients with FLG gene mutations are at an increased risk for developing asthma, but only in the context of having AD, pointing to the importance of allergic sensitization through a damaged skin barrier.44,47,48 Conversely, AD patients with more polarized Th2-type disease with allergies and asthma and increased biomarkers including serum IgE, TSLP and cutaneous T cell-attracting chemokine were also more likely to have severe skin disease complicated by eczema herpeticum (EH), Staphylococcus aureus or molluscum infections.49 In addition, patients with FLG mutations have been found to have an increased risk for EH, a serious complication of AD.46 Using a proteomics approach, Howell et al identified S100/A11 as a target in Th2 cytokine-mediated inhibition of FLG and the antimicrobial peptide, HBD-3, expression in AD skin, pointing to immune dysregulation effecting both epidermal barrier integrity and innate immune response.50 Still, the relationship of skin barrier and immune abnormalities to the increased susceptibility to microbial colonization and infections remains to be fully elucidated.34 Of interest, emerging observations that topical calcineurin inhibitors can in part correct the barrier defect in AD and that gentamicin can restore the production of functional FLG chains provides further evidence of the complex relationship of the epidermal barrier and the immune system.51
Effects of topical therapy on skin barrier in AD
It has been shown that skin barrier function as assessed by transepidermal water loss (TEWL) is intrinsically compromised in children with AD, but not in children with other allergic conditions.52 In addition, TEWL was higher in white than in African American children with AD and the magnitude of skin barrier dysfunction correlated with disease severity.52 While TEWL might be a useful biomarker in AD, racial and pigmentation differences will need to be considered. Recently, authors examined TEWL and other parameters of epidermal barrier and showed improvement in all parameters when AD patients were treated with both a topical steroid (betamethazone valerate) and a topical calcineurin inhibitor (pimecrolimus) applied to matched lesions.53 Both treatments normalized epidermal differentiation and reduced epidermal hyperproliferation. Betamethazone valerate was better in reducing clinical symptoms and epidermal proliferation, however it induced epidermal thinning. In contrast, pimecrolimus did not cause skin atrophy. Therefore, we could conclude that pimecrolimus may be used for long-term treatment, and betamethazone valerate might be more useful for treatment of acute exacerbations of AD.54
 XML Download
XML Download