

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Bladder cancer (BC) is one of the leading causes of cancer-related death worldwide, with an estimated 429,800 new cases and 165,100 deaths globally in 2012 [1]. BCs can be categorized into 2 groups: nonmuscle invasive BC (NMIBC) and muscle invasive BC (MIBC). Approximately 70% to 80% of patients are diagnosed with NMIBC, and some of these patients have a high risk of recurrence and a variable risk of progression despite administration of local therapies [2]. The remaining 25% of patients with MIBC are managed by radical surgery or radiotherapy, but often still have poor outcomes, despite administration of these systemic therapies [2]. For patients with advanced BC or failure of first-line chemotherapy, no clearly defined second-line treatments have been shown to prolong overall survival [2]. Therefore, elucidation of novel molecular mechanisms of BC progression and identification of novel tumor markers that can predict recurrence or survival are urgently needed.
Despite its relatively low sensitivity (30%–40%), urinary cytology is a reliable marker for BC diagnosis because of its high specificity (90%–95%). However, because of the low sensitivity of urinary cytology, patients are still forced to undergo a painful cystoscopic procedure to confirm the initial diagnosis and to check for recurrence during follow-up examinations after undergoing transurethral resection of bladder tumors. Several new urine-based tests for BC, such as bladder tumor antigen, nuclear matrix protein 22 (NMP22, Sysmex Corp., Kobe, Japan), and FISH (UroVysion, Abbott Molecular Inc, Des Plaines, IL, USA), have been approved for clinical use. However, these new urinary markers have not been widely applied because they have relatively low specificities (60%–80%) compared with urinary cytology, despite their higher sensitivities (50%–70%) [3]. Thus, specificity may come at the cost of sensitivity, and conventional urinary cytology is a good example of this compromise. Accordingly, new effective diagnostic markers are urgently needed for patients with BC.
MicroRNAs (miRNAs) are endogenous small noncoding RNA molecules (19–22 bases in length) that negatively regulate the expression of protein-coding genes in a sequence-specific manner [4]. The nature of miRNAs is unique in that one miRNA can regulate multiple protein-coding RNAs. Bioinformatics predictions have indicated that miRNAs regulate 30% to 60% of the protein-coding genes in the human genome [56]. Growing evidence has demonstrated that aberrantly expressed miRNAs can act as oncogenes or tumor suppressors in various types of malignancies [7]. These miRNAs can disrupt tightly controlled RNA networks in cancer cells [8]. Since 2006, we have rigorously investigated the molecular networks of miRNAs and their target genes in clinical BC. Identification of aberrantly expressed miRNAs in BC could provide important clues for the investigation of novel molecular mechanisms of initiation, progression, and metastasis in BC. In this review, we discuss the role of miRNAs in BC on the basis of up-to-date results from our laboratory and other research groups.
CURRENT CONSENSUS OF EPIGENETIC GENE REGULATION BY NONCODING RNA
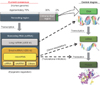
After the completion of the Human Genome Project in 2003, about 20,000 to 25,000 genes were identified, and the sequences of 3 billion base pairs of human DNA were determined [9]. Surprisingly, only 2% of the human genome encodes functional proteins; 50% to 75% of the human genome is transcribed, and 98% of the transcripts are not translated into protein. Fig. 1 shows the current consensus of epigenetic regulation by noncoding RNAs transcribed from approximately 70% of the genomic region that used to be considered as "junk DNA" [10]. These noncoding RNAs can be roughly classified into two groups according to size. The first group includes short RNAs of less than 200 nucleotides (nt) in length; miRNAs, which are small RNA molecules of around 18 to 22 nt in length, fit into this group, as do other classes of small RNAs, such as piwi-interacting RNAs (around 23–30 nt) [11]. The other group includes long noncoding RNAs (lncRNAs) of around 200 nt or more. lncRNAs have been implicated in a range of developmental processes and diseases and can inhibit or activate gene expression through a range of diverse mechanisms. In contrast to miRNAs, which have been extensively studied and have well-understood functions in gene regulation, lncRNAs are not well studied or characterized.
MICRORNAS BIOGENESIS
miRNAs are evolutionarily conserved and are located either within the introns or exons of protein-coding genes (70%) or in intergenic regions (30%). Most intronic and exonic miRNAs are derived from their host genes, which suggests that they are transcribed concurrently with their host transcripts. Transcripts containing primary miRNAs (pri-miRNAs), which can vary from 200 nt to several kb in length, are capped with a specially modified nucleotide at the 5'-terminus and are polyadenylated with multiple adenosines at the 3'-end. Pri-miRNA is cleaved into precursor-miRNA (pre-miRNA: 60–70 nt in length) by the RNAse III enzyme (also known as Drosha). Pre-miRNA is exported from the nucleus into the cytoplasm by exportin-5. In the cytoplasm, pre-miRNA is cleaved by another RNase III enzyme, known as Dicer, into miRNA duplexes of approximately 19–22 nt in length. One miRNA duplex is then recruited into the RNA-induced silencing complex and functions to recognize complementary sites within the target messenger RNA (mRNA), thereby regulating translation through mRNA cleavage, degradation, or transcriptional repression [12].
MICRORNA PROFILING STUDIES IN BLADDER CANCER
miRNA expression profiling by array-based and polymerase chain reaction (PCR)-based methods is not commonly used because these methods only allow for identification of a limited number of known miRNAs. The rapid development of high-throughput, deep sequencing (DS) technology has provided novel information regarding the functions and features of miRNAs. Han et al. [13] first reported miRNA profiling in BC by using an Illumina Genome Analyzer IIX in 2011. Soon thereafter, five studies used DS technology to establish the miRNA profiles of clinical BC samples [1415161718]. To date, the largest DS study on BC is a comprehensive investigation by The Cancer Genome Atlas Research Network [18]. Tissues from 131 patients with high-grade MIBC were analyzed by RNA sequencing together with 118 blood samples from the same individuals and 23 adjacent histologically proven normal tissues. The aim of the project was to carry out genetic and epigenetic characterization of the molecular landscape of high-grade MIBC. In our studies, we conducted miRNA profiling by use of both PCRbased methods and DS technology [1719]. When comparing our 2 studies, the altered expression of miRNAs detected by DS technology was very similar to the results of PCRbased methods. In addition, a considerable number of novel miRNAs have been reported by some studies [131417]. Therefore, this analysis supports that DS technology is superior to previously used methods. Although PCR-based and DS profiles are not comparable from a quantitative point of view, sequencing approaches should provide substantial improvements to transcriptome analyses.
MOLECULAR TARGETS OF MICRORNAS IN BLADDER CANCER
1. Features of miRNA studies in BC
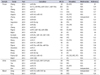
Growing evidence has demonstrated that aberrantly expressed miRNAs can act as oncogenic miRNAs (oncomiRNAs) or tumor-suppressive miRNAs (TS-miRNAs) in BC [1220]. Because miRNAs can construct miRNA-target gene networks that contribute to tumor initiation, survival, and invasion, many investigators have focused on the genes targeted by aberrantly expressed miRNAs in BC in comparison with normal bladder epithelium. In terms of miRNA analysis, luciferase-reporter assays are useful for confirming the direct binding of miRNAs to the specific 3'-untranslated region of the mRNA representing the target gene. Hence, we have summarized the miRNA studies in BC for which the target genes were validated by luciferase-reporter assays. To date, we have identified 78 studies that satisfied our criteria (Tables 1 and 2). As shown in Table 1, several TS-miRNAs, including miR-1, miR-16, miR-24, miR-101, miR-125b, miR-129, and miR-133a, have been shown to be located on different chromosomal regions but to have common sequences in their mature miRNAs. This suggests that the expression of these TS-miRNAs may be maintained by another genomic region, even though one region may be functionally disordered because of methylation, deletion, or mutation. This type of backup system is thought to have important roles in protecting normal cells from carcinogenesis.
The miRNAs in Table 1 are listed in order of miRNA number, and the validated target genes are indicated. Interestingly, among the 54 miRNAs examined, 45 were downregulated, whereas 9 were upregulated. In our experience, we often encountered difficulties with the use of anti-miRNAs in in vitro studies; however, miRNA restoration systems using miRNA transfection are well established and commercially available. Accordingly, the latter system may be easier to use to examine the potential tumor-suppressive roles of the downregulated miRNAs.
miRNAs have been shown to regulate target gene expression. Several miRNAs and their targets have been identified in BC (Table 2). Among the 61 target genes examined, the expression levels in clinical BC specimens compared with the levels in normal bladder epithelium were validated for 40 genes. The downregulation of several miRNAs, including miR-1, miR-133a, miR-145, miR-195, and the miR-200 family, has been reported, and these miRNAs have been shown to be involved in crucial cellular processes, such as the epithelial-mesenchymal transition (EMT), cell cycle, apoptosis, and cancer signaling pathways. TS-miRNAs and their target genes were classified into functional annotations by using the Kyoto Encyclopedia of Genes and Genomes pathways (Table 2).
2. Clustered miRNAs
Several miRNAs are located in close proximity (within 10 kbp) in the same genomic region; this is called a miRNA cluster. Because of their simultaneous expression, common target genes of miRNAs within a cluster may be important for tumor biology. For example, miR-145 forms a cluster with miR-143 on chromosome 5q32, and these clustered miRNAs are regulated by a common promoter [21]. Research indicates that both miR-145 and miR-143 are frequently downregulated in a broad range of cancers, including BC. Villadsen et al. [22] suggested that SERPINE1 is a direct target of both miR-145 and miR-143; this was the first report of an miRNA cluster targeting the same mRNA in BC. Importantly, miR-145 has been implicated in the p53 regulatory network in human malignancies [23]. p53 transcriptionally induces the expression of miR-145 by interaction with a potential p53 response element in the miR-145 promoter; moreover, MYC is directly repressed by miR-145. Additionally, Chiyomaru et al. [24] found direct binding of the miR-23b/27b cluster with EGFR/MET, and Itesako et al. [17] found direct binding of the miR-195/497 cluster with BIRC5, which encodes survivin.
3. Cell cycle/apoptosis-related genes
Avoidance of apoptosis and cell cycle acceleration are required for cancer cells to escape cell death. BCL2L2, a member of the BCL2 family, can promote cell survival by inhibiting the intrinsic pathway of apoptosis. In contrast, BIRC5 (survivin) is a key member of the inhibitor of apoptosis protein family. Several TS-miRNAs induce apoptosis through targeting BCL2L2 (miR-133b, miR-203) and BIRC5 (miR-195, miR-203, miR-497). Upregulation of miR-129 may be involved in avoidance of apoptosis through direct targeting of SOX4 and GALNT1, which are involved in transcription and protein expression, respectively [25]. One important pathway is the sequential activation of caspases, which have a central role in the execution of cell apoptosis. miR-1 is downregulated in both clinical BC tissues and cultured BC cells and has been shown to induce apoptosis by increasing the activity of caspases 3 and 7 through direct targeting of the apoptosis inhibitor SRSF9 [26]. Additionally, CASP2 (caspase 2) is directly targeted by an onco-miRNA, miR-706 [27].
Several miRNAs have been shown to control the cell cycle through their target genes. Matsushita et al. [28] first reported that cell-cycle-related genes, such as CCNE1/2, CDC25A, and PKMYT1, are directly regulated by miR-144-5p. CDC25A is also regulated by miR-449a, and CDK4 and CDK6 are regulated by miR-124-3p, miR-195, miR-29c, miR-230c, and miR-449a, inducing G1-phase arrest. Interestingly, miR-96, which is upregulated in BC, directly binds to CDKN1A (p21), a well-known tumor-suppressive gene [29]. Moreover, miR-125b and miR-200c target the oncogene E2F3, which is critical for the G1/S transition and is overexpressed in most high-grade BCs [3031]. Inoguchi et al. [32] found that miR-24 regulates FOXM1, which is critical for mediating cell cycle progression proteins, such as Aurora B, cyclin B1, and CDC25B, and stimulates the expression of SKP2/CKS1, which is involved in the proteolysis of p27kip1 and in G1/S progression.
4. EMT-related genes as targets of miRNAs in BC
We also evaluated whether the expression of the different target genes could be validated in clinical BC samples (Table 2). The EMT is a key progression mechanism that is often activated during cancer invasion and metastasis and is characterized by loss of cell adhesion, repression of E cadherin expression, and increased cell mobility [33]. Members of the miR-200 family are well-known regulators of the EMT in human malignancies, including BC [34], and some investigators have reported the functions of these miRNAs in BC [3536].
FSCN1 and LASP1 are components of the filopodia and lamellipodia in various types of cells; both proteins are involved in mediating the dynamics of actin filaments and are activated during the EMT. miR-133a and miR-145 directly regulate FSCN1 [37], and the miR-1/133a cluster and miR-218 regulate LASP1 [38]. Restoration of the expression of these miRNAs accelerates cell migration and invasion activity in vitro [3738]. Matrix metalloproteinases (MMPs) are calcium-dependent zinc-containing endopeptidases that are capable of degrading various extracellular matrix proteins. MMPs are involved in the cleavage of cell surface receptors, the release of apoptotic ligands (such as the FAS ligand), and chemokine/cytokine inactivation. MMPs are also thought to facilitate tumor cell invasion and metastasis via the degradation of the extracellular matrix. Transforming growth factor beta1 reduces the expression level of miR-200b, which directly regulates MMP16. This then activates other MMPs (e.g., MMP-2 and MMP-9), growth factors, and receptors, thereby facilitating local cellular mechanisms that promote migration [39].
Loss of expression of miR-23b, miR-138, and miR-200c (an miR-200 family member) is associated with increased expression of ZEB1, ZEB2, and ERFFI1, which facilitate the progression of the EMT [364041]. ZEB1 and ZEB2 are known to negatively regulate E cadherin expression and are directly targeted by miR-23b and miR-138, whereas ERRFI-1, a regulator of EGFR, is directly targeted by members of the miR-200 family [36]. ZEB1 expression has been reported in 22% of clinical BC tissue specimens, but is absent from the bladder mucosa [42]. Expression of the miR-200 family has been shown to be repressed by hypermethylation of its promoter region, and loss of miR-200c expression is significantly associated with subsequent disease progression to MIBC and poor outcomes [35]. These studies implied that many EMT-related genes are activated through downregulation of several TS-miRNAs, and these phenomena may contribute to tumor progression of BC.
5. Others
Downregulated miRNAs have also been reported to target some genes related to cytokine–cytokine receptor interactions, including EGFR (miR-23b, miR-27b, miR-133a, miR-133b) [2443], IGF1R (miR-145) [44], and MET (miR-23b, miR-27b, miR-101) [2443]. Activation of these genes may affect downstream cancer pathways, such as mammalian target of rapamycin signaling, phosphoinositol 3-kinase (PI3K)-Akt signaling, and mitogen-activated protein kinase (MAPK) signaling. FGFR3, which belongs to the MAPK signaling pathway, is crucial for BC oncogenesis, and mutations in FGFR3 have been shown to accelerate proliferation in BC [45]. Downregulation of miR-99a and miR-100 has been shown to cause upregulation of FGFR3 expression before its mutation, which suggests that the acquisition of mutations may result from increased cell turnover [46]. The expression of AKT1 and ILK, which are involved in the AKT-PI3K pathway, was shown to be repressed by the miR-143/145 cluster, and proliferation is inhibited in cells transfected with these miRNAs [4748]. The vascular endothelial growth factor (VEGF) and WNT signaling pathways are also important for BC development, and VEGFC, FZD4, PSEN1, and WNT1 have been reported to be under the control of several TS-miRNAs [1749505152].
MICRORNAS AS TUMOR MARKERS IN BLADDER CANCER
1. miRNAs as diagnostic markers for BC detection
Many studies have investigated miRNA expression by quantitative real-time PCR of clinical BC specimens [20]. In this review, we focus on studies in which the investigators evaluated diagnostic accuracy by use of receiver-operator curve analyses of the sensitivity and specificity of specific miRNAs in comparison with normal bladder epithelium. We found 18 studies that satisfied our criteria (Table 3). In 2009, Ichimi et al. [19] first evaluated the expression of six miRNAs in tissues as diagnostic markers to distinguish BC (n=104) from non-BC (n=31).
There is a pressing need for highly accurate, noninvasive tests for BC diagnosis. The standard procedures for BC diagnosis and the follow-up for patients depend on urinary cytology and cystoscopy. Urinary cytology is a reliable urine marker for BC diagnosis because of its high specificity (90%–95%). However, this method has low sensitivity (30%–40%), and patients are forced to undergo a painful cystoscopic procedure to confirm the diagnosis. A number of noninvasive urine tests, such as bladder tumor antigen and NMP22, have been developed; however, their diagnostic specificities are inferior to that of urinary cytology tests, and these methods have failed to meet clinical expectations. Therefore, quantitative changes in miRNAs in urine, blood, and tissues are the primary focus in the search for new markers. miRNAs are detectable in urine with pelleted or cell-free urine samples. Hanke et al. [53] evaluated miR-126 expression in whole urine samples and showed that this miRNA had a sensitivity of 72% and specificity of 82% when used as a diagnostic marker, even though the sample size in their study was small. Yamada et al. [54] also found that miR-96 detection in the urine including the pellet was a useful diagnostic marker, with a sensitivity of 71.8% and a specificity of 89.2%. Shimizu et al. [55] uniquely reported good sensitivity of 81% and good specificity of 89% by testing a methylation panel of miR-9-3/miR-124-2/miR-124-3/miR-137. In terms of miRNA detection in cell-free urine [5657585960], several studies have shown that this method provides moderate sensitivity and specificity for distinguishing BC from non-BC but that the results are inferior to those from studies using whole urine samples [5354556162]. In blood tests, Jiang et al. [63] showed dramatic results (sensitivity of 90% and specificity of 90%) using a panel study of miR-15b-5p/miR-27a-3p/miR-30a-5p/miR-148b-3p/miR-152/miR-3187-3p.
2. miRNAs for predicting prognosis in patients with BC
Many retrospective studies have suggested that the expression of specific miRNAs in BC specimens may be a good prognostic marker predicting overall survival, disease-free survival, recurrence-free survival, or progression-free survival, as outlined in Table 4. Moreover, several reports analyzing urine and blood samples have shown that miR-200, miR-214, and miR-3187-3p levels are good prognostic markers in the blood [57606364]. Surprisingly, miR-203, miR-214, miR-152, and miR-3187-3p were shown to be independent markers predicting overall survival, progression-free survival, or recurrence-free survival in multivariate studies [636465]. Unfortunately, no prospective studies have been performed to date. Thus, additional large, prospective studies are needed for the clinical application of miRNA assays for diagnostic and prognostic purposes in BC.
MICRORNAS AS A TREATMENT MODALITY IN BLADDER CANCER
Some TS-miRNAs have been transfected into BC cell lines by using plasmid/virus vectors, followed by subcutaneous injection of the transfected cells into nude mice [276667]. Significant inhibition of tumor growth [2767] and metastasis [66] was observed. Yu et al. [66] found that the CD44 gene was a direct target of miR-34a and that the pro-apoptotic role of miR-34a may be mediated primarily through regulation of CD44, which functions to promote metastasis and angiogenesis. Xu et al. [68] demonstrated that miR-100-transfected BC cells exhibited significant growth inhibition in an intravesical orthotopic BC model. In terms of onco-miRNAs, miR-10b- and miR-155-transfected BC xenografts exhibited significantly increased growth in comparison with controls [6970]. Inamoto et al. [71] tested the anticancer effects of intravesical injection of miR-145 by using cationic liposomes (Lipofectamine RNAiMAX, Thermo Fisher Scientific, Waltham, MA, USA) in an intravesical orthotopic BC model. They achieved adequate anticancer effects after intravesical administration of miR-145. These miRNAs are promising candidate miRNAs that may be useful for clinical applications in the future.
FUTURE PERSPECTIVES
Many studies have examined the roles of miRNAs in human malignancies, including BC. However, the majority of miRNA studies have focused on candidate gene approaches with limited whole-genome sequencing, and a detailed characterization of miRNA profiles based on the genomic features of BC has not been reported. Recently, DS technology has been applied for profiling of mRNA and miRNA in human cancers, including BC. However, DS technology has mainly focused on BC tissues without any particular selection according to clinical treatment. Additional DS analyses should be conducted using samples from patients who have undergone different treatments, e.g., chemotherapy regimens, radiation therapies, or immune-checkpoint drugs.
A growing body of evidence has shown that miRNAs have great potential for clinical use as new diagnostic biomarkers. However, detection of single miRNAs in urine samples will not be superior to current urinary cytology tests because of the relatively low diagnostic specificity of such analyses. Some recent studies have attempted to improve the diagnostic specificity of miRNA detection by using a panel of multiple miRNAs [55566163]. It will be important to select optimal combinations of miRNAs and to perform prospective translational studies in outpatients to validate such miRNA panels.
Moreover, recent studies have shown that some miRNAs control the activity of major cancer-related signaling molecules. Thus, identification of aberrant miRNA expression and oncogenic or tumor-suppressive molecular targets of miRNAs is necessary for the clinical development of novel cancer therapeutics. Because many TS-miRNAs and their target oncogenes are components of complex molecular networks, treatment with multiple miRNAs may provide stronger anticancer effects than treatment with a single miRNA. In an interesting study by Liu et al. [72], new vectors, termed "miRNA-mowers," that contained the entire sequence of the onco-miR-183-96-182 cluster were constructed. Transfection of the miRNA-mowers strongly inhibited cell growth and migration and induced apoptosis in vitro, suggesting the usefulness of targeting multiple onco-miRNAs. However, no studies have yet reported successful results for intravenous administration of TS-miRNAs in the induction of anticancer effects in vivo. The development of an adequate drug delivery system for TS-miRNAs is needed; otherwise, it will be difficult to determine the value of miRNAs as a therapeutic modality.



 XML Download
XML Download