

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Cordyceps militaris is also known as the rare Chinese caterpillar fungus (1) and has benefits for the human body including effects on the circulatory, immune, respiratory, and glandular systems. It is commonly used in the orient to replenish the kidneys and soothe the lung as well as in the treatment of hyperglycemia, hyperlipidemia (2,3), renal dysfunction, and liver disease (4); it is also thought to have anti-mutagenic (5) and anti-angiogenesis (6) capabilities. Recently, several studies demonstrated that extracts of Cordyceps militaris had multiple pharmacological actions which included anti-inflammatory activity (6), improvement of insulin resistance and insulin secretion (2), and tumor suppression activity. Addition, it possesses antioxidant activity to a greater extent than Cordyceps sinensis or Cordyceps kyushuensis (7). Because natural Cordyceps militaris is rare and expensive, many scientists have examined its life cycle with the aim of developing techniques for the isolation of fermentable strains. Even though the immunomodulatory role of CME in immune response is being explored and further elucidated, the regulatory effects of CME on specific functions of macrophages has yet to be fully explained.
Macrophages, a type of differentiated tissue cell which originate as blood monocytes, play an important role in immune and allergic reactions as well as in inflammation (8). These cells have several functions including the killing and removal of pathogenic microbes and the processing and presentation of antigens which have been ingested by lymphocytes (9). Macrophages are essential cellular components of the body's host defense system, playing critical roles in both innate and adaptive immunity. Also, these cells collaborate extensively with T cells through cell-cell and cytokine-mediated interactions to coordinate the evolution of inflammatory responses. In response to microbes and their products, such as lipopolysaccharide (LPS), macrophages secrete various inflammatory cytokines including interleukin (IL)-1β, IL-6, IL-12, and tumor necrosis factor (TNF)-α through the activation of nuclear factor (NF)-κB; they also express NF-κB-dependent inducible nitric oxide (NO) synthase (iNOS) (10,11) and cyclooxygenase-2 (COX-2) (12). Many diseases, such as rheumatoid arthritis (RA), arteriosclerosis, chronic hepatitis, and pulmonary fibrosis (13-16) involve the overproduction of inflammatory mediators. Although large amounts of macrophage-derived proinflammatory mediators can cause several immune diseases, proper regulation of macrophage function by immunomodulating agents could help protect the host from various pathogenic and cancerous attacks.
Nuclear factor-κB, a nuclear transcription factor, regulates the expression of various genes, including IL-1β, TNF-α, iNOS, and COX-2, that play critical roles in apoptosis, tumorigenesis, various autoimmune diseases, and inflammation (17,18). Because of its ubiquitous role in the pathogenesis of inflammatory gene expression, NF-κB is currently a target for treating various diseases (19). Nuclear factor-κB is activated in response to various inflammatory stimuli, including bacterial LPS, cytokines, and viral proteins (20). Under normal conditions, NF-κB is present in the cytoplasm as an inactive hetero-trimer consisting of p50, p65, and IκBα subunits. Upon activation, degradation of IκBα exposes a localization signal on the p50/p65 hetero-dimer leading to nuclear translocation and to its binding to a specific sequence in several promoters which, in turn results in the transcription of proinflammatory genes.
Given that these effects have been suggested to play an important role in disease therapy, we investigated whether CME could increase the production of proinflammatory mediators in macrophages.
MATERIALS AND METHODS
Reagents
Lipopolysaccharide (LPS) was purchased from Sigma (St. Louis, USA). Cell culture media DMEM, antibiotic-penicillin/streptomycin solution, and fetal bovine serum (Hyclone, Logan, USA) were used for the cell culture.
Preparation of CME from Cordyceps militaris
Cultures of fruiting bodies of Cordyceps militaris were supplied by CM Biotec, Kangnung, Korea and identified by Professor Jae-Mo Sung (Department of Agricultural Biology of Kangwon National University). The dried cultured fruit bodies were extracted with distilled deionized water (DDW) for 6 hrs at 60℃. The water extracts were concentrated with a vacuum evaporator at 70℃ then lyophilized. Cordyceps militaris extract (CME) was included the cordycepin (3'-deoxyadenosine) 1.3 ng/mg which was purchased from Sigma (St. Louis. MO, USA).
Cell culture
RAW264.7 mouse macrophage cells (American Type Culture Collection) were maintained in Dulbecco's Modified Eagle's Medium (DMEM), supplemented with 10% heat-inactivated fetal bovine serum (FBS; Hyclone, USA), 100 U/ml of penicillin, and 100 g/ml of streptomycin (Hyclone, USA) at 37℃ in a 5% CO2 incubator. Primary macrophages were collected from the peritoneal cavities of mice (8-week-old male ICR mice) after an intraperitoneal (i.p.) injection of 3 ml of 3% thioglycolate broth (Sigma, St. Louis, MO) 4 days before harvesting. The peritoneal macrophages were then washed with 1×PBS (Ca2+- and Mg2+-free) and plated with DMEM containing 10% FBS, 100 U/ml of penicillin, and 100 U/ml of streptomycin at 37℃ in a 5% CO2 incubator.
MTT assay for cell viability
A commercially-available cell viability assay was employed to evaluate the cytotoxic effect of cordycepin using thiazolyl blue tetrazolium bromide (Sigma, St. Louis, USA). RAW264.7 cells (2×105 cells/well) were plated with various concentrations of CME in 96-well microtiter plates (Nunc, Roskilde, Denmark) and were then cultured overnight at 37℃ in a 5% CO2 incubator. Afterwards, 50 µl of MTT solution was added to each well, and the cells were then cultured for 4 hrs at 37℃ in a 5% CO2 incubator. 100 µL of solubilized solution were added to each well. The plate was allowed to stand overnight in the incubator after evaluation for complete solubilization of the purple formazan crystals and measurement of the optical density (OD) at 560 nm by a microplate reader (Molecular Devices corporation, Sunnyvale, USA).
Measurement of NO and PGE2
The amount of nitrite and PGE2 produced by mouse macrophages was measured in cell culture supernatant. The cells were plated at a density of 1×106 cells in a flat-bottomed, 96-well plates with the 200 µl of culture medium. They were then treated with various concentrations (12.5~200 µg/ml) of CME in the absence of LPS (10 ng/ml) and incubated for overnight. NO production was determined according to the method reported by Stuehr and Nathan (21). The amount of nitrite was measured using the Griess reagent (stock-I: 0.2% N-(1-naphthyl) ethylenediamine-HCl, stock-II: 2% sulfanilamide in 5% H2PO4) and the amount of PGE2 produced was measured using an enzyme-linked immunosorbent assay (ELISA) kit (R&D system, Minneapolis, MI, USA), according to the manufacturer's instructions.
Cytokine assays
The amount of TNF-α, IL-1β, and IL-6 in the cell culture supernatant were measured using an ELISA kit (eBioscience, San Diego, CA, USA). RAW 264.7 cells were cultured in DMEM with 10% FBS in 12-well, flat-bottomed plates at a density of 5×105 cells/well. The cells were treated with various concentrations (12.5, 25, 50, 100, 200 µg/ml) of CME in the absence or presence of LPS (50 ng/ml) at 37℃ for 48 hrs in humidified air with 5% CO2. Subsequently, the culture supernatant was collected and assayed according to the manufacturer's instructions.
Isolation of total RNA and RT-PCR
Total RNA was extracted from RAW264.7 cells using the RNeasy Mini kit (QIAGEN, USA) in an RNase-free environment. RNA was quantified by reading the absorbance at 260 nm as previously described (14). The reverse transcription of 1 µg RNA was carried out using M-MLV reverse transcriptase (Promega, USA), oligo (dT) 16 primer, dNTP (0.5 µM), and 1 U RNase inhibitor. After incubation at 65℃ for 5 min and 37℃ for 60 min, M-MLV reverse transcriptase was inactivated by heating at 70℃ for 15 min. The polymerase chain reaction (PCR) was performed in 50 mM KCl, 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, and 2.5 mM dNTPs with 5 units of Taq DNA polymerase and 10 pM of each primer set for iNOS, PGE2, IL-1β, IL-6, and PGE2. The cDNA was amplified by 35 cycles of denaturing at 94℃ for 45 s, annealing at 62℃ for 45 s, and extension at 72℃ for 1 min. Final extension was performed at 72℃ for 5 min. The PCR products were electrophoresed on a 1.5% agarose gels and stained with ethidium bromide. The primers used were 5' AGCTCCTCCCA GGACCACAC 3' (forward) and 5' ACGCTGAGTACCTCATTGGC 3' (reverse) for iNOS, 5' CAGGATGAGACATGACACC 3' (forward) and 5' CTCTGCAGACTCAAACTCCAC 3' (reverse) for IL-1β, 5' GTACTCCAGAAGACCAGAGG 3' (forward) and 5' TGCTGGTGACAACCACGGCC 3' (reverse) for IL-6, 5' TTGACCTCAGCGCTGAGTTG 3' (forward) and 5' CCTGTAGCCCACGTCGTAGC 3' (reverse) for TNF-α, 5' AAGAAGAAAGTTCATTCCTGATCCC 3' (forward) and 5' TGACTGTGGGAGGATACATCTCTC 3' (reverse) for COX-2, and 5' GTGGGCCGCCCTAGGACCAG 3' (forward) and 5' GGAGGAAGAGGATGCGGCAGT 3' (reverse) for β-actin. β-actin was used as an internal control.
Preparation of nuclear extracts
After cultured cells were collected and washed twice with cold PBS, they were resuspended in hypotonic buffer (10 mM HEPES, pH 7.9, 10 mM KCl, 1.5 mM MgCl2, 0.2 mM PMSF, 0.5 mM DTT, 10 µg/ml aportinin). After 15 min incubation on ice, the cells were lysed by the addition of 0.1% NP-40 and vigorous vortexing for 1 min. The nuclei were pelleted by centrifugation at 12,000×g for 1 min at 4℃ and resuspended in high salt buffer (20 mM HEPES, pH 7.9, 25% glycerol, 400 mM KCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM DTT, 1 mM NaF, 1 mM sodium orthovanadate). The supernatant fluid was stored in aliquots at -70℃.
Western blot analysis
RAW 264.7 cells were washed with phosphate-buffered saline (PBS) and lysed by lysis buffer (1% SDS, 1.0 mM sodium vanadate, 10 mM Tris-Cl buffer, pH 7.4) for 5 min. 20 µg of protein from the cell lysates was applied to 8~12% SDS-polyacrylamide gels and then transferred to nitrocellulose membranes. The membranes were blocked with 5% skim milk in PBST solution for 1 hr. They were then incubated with anti-IL-1β, anti-IL-6, anti-TNF-α, anti-i-NOS, anti-COX-2, and anti-NF-κB monoclonal antibodies (Santa Cruz Biotechnology Inc., Santa Cruz, California, USA) for 2 hrs and washed 3 times with PBST. After incubation with an alkaline phosphatase-labeled secondary antibody (Abcam, Cambridge, USA) for 2 hrs, the bands were visualized using a Western Blot Kit with alkaline phosphatase substrate (Vector, Burlingame, USA).
Flow cytometry
RAW 264.7 cells (1×106 cells/ml) were cultured in Petridishes. The cells were treated with various concentrations (12.5, 50, 200 µg/ml) of CME in the absence or presence of LPS (100 ng/ml). the dishes were incubated at 37℃ for 24 hrs in humidified 5% CO2 incubator under standard conditions. The cells were then washed with PBS. The washed cells were blocked with staining buffer containing 10% normal mouse serum (NMS) for 20 min on ice. The blocked cells were incubated with co-stimulatory molecules such as ICAM-1, B7-1 and B7-2 antibody (BD Biosciences, San Jose, USA) for 20 min on ice. The incubated cells were washed three times with staining buffer and then fixed by 1% paraformaldehyde in PBS. The fixed cells were measured by flow cytometry (Beckman coulter, Brea, USA).
Data analysis
Data are expressed as mean±standard deviation. Statistical significance between the groups was determined by a paired t-test and one-way ANOVA for repeated measures. Results of p<0.05 were considered statistically significant. Data were assessed using SPSS (version 15.0, SPSS Inc., Chicago, Illinois).
RESULTS
CME enhances NO and PGE2 production in macrophages
To analyze the potential proinflammatory properties of CME, we used murine macrophages, which produces NO and PGE2. Nitric oxide is known to represent toxic and proinflammatory mediators in acute and chronic inflammatory diseases as well as in normal defense reactions. In the current study, macrophages did not release NO in response to the medium alone; LPS (10 ng/ml) was used as a positive control for macrophage activation (Fig. 1A and B). When various concentrations of CME (12.5, 25, 50, 100, 200 µg/ml) were added to the culture media at the time of cell stimulation (18 hrs), NO production increased in a CME concentration-dependent manner (Fig. 1A and B). Further, CME was also found to dose-dependently increase PGE2 production (Fig. 1C and D). The potential cytoxicity of CME was evaluated by MTT assay after incubating cells for 24 hrs in the absence or presence of LPS. Results revealed that viability was unaffected at the concentrations used to enhance NO and PGE2 (data not shown). Thus, the immune enhancement was not attributable to a cytotoxic effect.
CME regulates the mRNA and protein expression of iNOS and COX-2
RT-PCR and western blot analyses were performed to determine whether CME had a direct effect on proinflammatory mediators (NO and PGE2) related to modulation of the expression of iNOS and COX-2. As shown in Fig. 2A, iNOS and COX-2 protein expression was markedly induced in macrophage cells after treatment both with CME and LPS. This induction was increased by CME treatment in a dose-dependently manner. Furthermore, RT-PCR analysis revealed that the expression of the COX-2 gene was correlated with their protein levels (Fig. 2B).
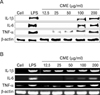
Direct cytokines production in response to CME
To determine whether CME had a direct effect on cytokine production, IL-1β, IL-6, and TNF-α secretion were measured in the macrophage cell line using cytokine ELISA kits. IL-1β, IL-6, and TNF-α, are the major proinflammatory cytokines produced by monocytes and macrophages. As shown in Fig. 3, CME dose-dependently increased cytokine production. Interestingly, CME also strongly increased IL-1β and IL-6 production.
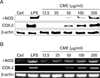
CME regulates the gene and protein expression of proinflammatory cytokines
To further investigate the important role of CME on immune response, inducing RAW 264.7 cells for 24 hours with LPS or CME resulted in the production of proinflammatory cytokines. As shown in Fig. 4A, CME concentration-dependently induced the expression of IL-1β, IL-6, and TNF-α protein and mRNA (Fig. 4B).
CME strongly modulates the surface expression levels of co-stimulatory molecules
Further to evaluate whether CME influences the phenotypic maturation of macrophages were cultured with CME or LPS for 24 h and then analyzed for any surface expression of ICAM-1, B7-1, and B7-2 molecules. As shown in Fig. 5, CME similarly to LPS, induced dramatic up-regulation of ICAM-1 and B7-1/-2 molecules.
CME induces of NF-κB activation
It has been shown that NF-κB activation is a critical factor for the expression of various cytokines, iNOS, and COX-2 in macrophages in response to LPS. In an unstimulated cell, NF-κB resides in the cytoplasm as an inactive NF-κB/IκB complex. Under stimuli, IκBα is phosphorylated and subsequently degraded, allowing NF-κB to translocate into the nucleus (13). To investigate whether CME could affect nuclear translocation of NF-κB, a western blot analysis for NF-κB p65 was carried out with cell lysate in macrophages (Fig. 6). The amount of NF-κB p65 markedly increased upon exposure to LPS alone; CME also increased NF-κB p65.
DISCUSSION
The data presented in this paper indicated that CME can exert significant immunomodulatory effects on macrophage-mediated immune responses. The present study demonstrated that CME is an effective activator of NO generation, cytokine (IL-1β, IL-6, and TNF-α) expression, PGE2 secretion, and co-stimulatory molecules in macrophages. Inflammation is a complex process involving numerous mediators of cellular and plasma origins. Activated macrophages fuse their lysosomes more efficiently to phagosomes, exposing intracellular or recently ingested extracellular microbes to a variety of microbiocidal lysosomal enzymes. Activated macrophages also produce oxygen radicals and NO, both of which have potent antimicrobial activity. Furthermore, they synthesize anti-microbial peptides and proteases that can be released to attract extracellular parasites. Additional changes in activated macrophages assist in amplifying the immune response.
Nitric oxide has been known to play a critical role in prevention against microbial infection and cancer occurrence (23,24). In addition, NO may be involved with regulation of apoptosis (25) and progression of autoimmune disease by altering the differentiation of the helper T cell phenotype (26). The biological activity of NO has become more complicated since NO production was reported to suppress the functional activity of T lymphocytes and antigen presenting cells (APCs) (27). In this paper, we have demonstrated that CME induced an increase of NO production in vitro (Fig. 1 and 2).
Prostaglandins, which are produced by COX-2, have been implicated as important mediators in endotoxemia and inflammatory conditions (28). For example, PGE2, like NO, is a pleiotropic mediator produced at inflammatory sites by COX-2 that gives rise to pain, swelling and stiffness (29). Here, we have demonstrated that CME significantly increases the gene expression of COX-2 and PGE2 production in macrophage cells (Fig. 1 and 2).
Cytokines are local protein mediators and are involved in almost all important biological processes including cell growth and activation, inflammation, immunity, and differentiation. In this study, CME dose-dependently enhanced IL-1β, IL-6 and TNF-α production in RAW 264.7 cells (Fig. 3 and 4).
The expression of cytokines requires the activation of NF-κB, a nuclear transcription factor, which regulates the expression of various genes, including IL-1β, i-NOS, and COX-2 all of which play critical roles in apoptosis and autoimmune disease. Nuclear factor-κB requires the phosphorylation of IκB and then targets IκB for ubiquitination and degradation. As shown in Fig. 6, CME increased proinflammatory mediators via NF-κB activation in RAW 264.7 cells.
The intracellular adhesion molecules (ICAMs), ICAM-1, ICAM-2 and ICAM-3, are cell surface ligands for leukocyte integrins. They are crucial in the binding of lymphocytes and other leukocytes to certain cells including APCs. In the current study, CME induced the ICAM-1 molecules in macrophages (Fig. 5A). The B7 family plays an important role as a co-stimulatory factor in APCs. Treatment with CME had another major effect on co-stimulatory molecules B7-1/-2 by strongly upregulating the surface levels of B7-1/-2 molecules in macrophage cells (Fig. 5B, C). All these molecules have been described to be of major importance in APC function (30,31).
In summary, the finding of the present study suggest that CME isolated from the fruiting bodies of Cordyceps militaris is a potent activator of NO, PGE2, and proinflammation production in macrophages, and that it acts at the level of transcription. Moreover, these inductions of the immune reaction by CME were associated with NF-κB activation. In conclusion, CME has immunostimulatory effects on macrophages through an enhancement of the production of cytokines and other bioactive substances as well as through the increased expression of co-stimulatory and adhesion molecules. In conclusion, together with these facts of our finding suggest that CME might be useful as a therapeutic agent for the treatment of the immune deficiency diseases.




 XML Download
XML Download