

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Selenoprotein P (SeP, SEPP1) is a glycoprotein that is mainly expressed in the liver and detected in plasma. SEPP1 plays a role in the transfer of selenium from the liver to plasma and other tissues. SEPP1 transcription is stimulated by interactions among the transcription factors forkhead box protein O1 (FoxO1), hepatocyte nuclear factor 4 (HNF4) α, and peroxisome proliferator-activated receptor-γ coactivator (PGC)-1α, which stimulate hepatic gluconeogenic gene expression. On the other hand, SEPP1 is attenuated by insulin [12]. Serum and hepatic SEPP1 levels reportedly have a strong positive relationship with the development of metabolic diseases [345]. In both animal and clinical studies, SEPP1 has been shown to induce insulin resistance due to dysregulation of insulin signaling and glucose metabolism in the liver and skeletal muscle [3]. Moreover, serum SEPP1 levels are inversely related to blood adiponectin levels in patients with type 2 diabetes, whereas increased adiponectin levels have been observed in SEPP1-knockout mice [6].
Fetuin-A (α2-Heremans-Schmid glycoprotein), a 64-kDA circulating liver-derived glycoprotein, serves as a biomarker for insulin resistance, nonalcoholic fatty liver disease (NAFLD), and cardiovascular disease [789]. Fetuin-A levels are high in human hepatocytes exposed to high levels of palmitic acid (PA), glucose, or endoplasmic reticulum (ER) stress activator thapsigargin [10]. Fetuin-A inhibits insulin-induced IRS-1 tyrosine phosphorylation, thus aggravating insulin resistance [11]. Moreover, fetuin-A-knockout mice exhibit enhanced glucose clearance, improved insulin sensitivity, and resistance to dietary fat-induced weight gain [12].
Exendin-4 (Ex-4), a potent glucagon-like peptide-1 (GLP-1) receptor agonist, is an incretin mimetic capable of relieving insulin resistance. It can decrease plasma glucose and triglyceride levels and increase high density lipoprotein cholesterol levels. The insulin-sensitizing effect of exendin-4 has been reported in human and animal models of insulin resistance [1314]. Exendin-4 also improves metabolic syndrome via modulation of the production and release of various cytokines associated in insulin resistance, oxidative stress, apoptosis, and inflammation [15]. GLP-1 analogue reportedly increases the level of the hepatokine fibroblast growth factor 21, which is involved in insulin sensitivity and glucose and lipid homeostasis [16]. However, the effect of exendin-4 on the expression of the novel hepatokines SEPP1 and fetuin-A remains unknown. Therefore, we evaluated changes in the expression levels of SEPP1 and fetuin-A after administration of exendin-4 under ER stress.
METHODS
Chemical reagents
PA, tunicamycin (Tuni), exendin-4, tauroursodeoxycholic acid (TUDCA), and 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Antibodies to fetuin-A, inositol-requiring enzyme-1α (IRE1α), CCAAT/enhancer binding homologous protein (CHOP), and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA). SEPP1 and phosphor-IRE1α (P-IRE1α) antibodies were purchased from Abcam (Cambridge, MA, USA). PKR-like endoplasmic reticulum kinase (PERK), phosphor-PERK (P-PERK), activating transcription factor 6 (ATF6), X-box binding protein 1 (XBP-1) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture and transfection
The HepG2 human hepatoma cell line (ATCC, Manassas, VA, USA) was cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum and 1% penicillin/streptomycin (Gibco, Grand Island, NY, USA). At approximately 85% confluence in six-well plates, cells were exposed to the ER stress inducers PA and Tuni. After 24 hours, cultured cells were treated with 100 nM exendin-4, 200 µM TUDCA, and 1 mM AICAR, after which they were incubated for 24 hours.
For gene knockout, the cells were transfected with 10 nM small interfering RNA (siRNA) of AMP-activated protein kinase (AMPK) and scrambled control siRNA (Santa Cruz) using the lipofectamine RNAiMAX reagent according to the manufacturer's instructions (Invitrogen, Carlsbad, CA, USA). After incubation for 24 hours, the transfected cells were treated with exendin-4 (100 nM) for 24 hours.
Total RNA isolation and real-time reverse transcription polymerase chain reaction
Total RNA was isolated from the cells using Trizol reagent (Invitrogen) to measure the messenger RNA (mRNA) levels of the SEPP1 and fetuin-A genes. Total RNA (2 µg) was reverse-transcribed to complementary DNA using the High Capacity RNA-to-cDNA Kit (Applied Biosystems, Foster City, CA, USA). After cDNA synthesis, quantitative real-time PCR was performed using SYBR green (Roche, Lewis, UK) and specific primers (Bioneer Co., Daejeon, Korea) according to the manufacturers' instructions. To normalize the expression of the target genes, the expression of β-actin (Actb) was used as an endogenous control in the comparative Ct method (2-delta delta Ct).
Western blot analysis
Cells were lysed in ice-cold RIPA buffer (Cell Signaling Technology) containing protease inhibitor cocktail (Roche) and phosphatase inhibitor cocktails (Sigma-Aldrich). The lysates were incubated on ice for 30 minutes, and then centrifuged at 13,000 rpm for 20 minutes at 4℃. The protein concentration of the supernatant was quantified using the Bradford protein assay (Bio-Rad Protein Assay, BioRad, Hercules, CA, USA) with bovine serum albumin standard (Thermo Scientific, Rockford, IL, USA). Equal amounts of protein (20 µg) were electrophoresed on 4% to 12% Bis-Tris Nupage gels (Invitrogen) and transferred to polyvinylidene difluoride membranes using the iBlot Dry Blotting System (Invitrogen). After transfer, the membranes were blocked in 5% bovine serum albumin /Tris buffered saline with Tween-20 buffer for 1 hour, and then incubated overnight at 4℃ with antibodies to SEPP1, fetuin-A, IRE1α, P-IRE1α, PERK, P-PERK, ATF6, XBP-1, CHOP, and β-actin followed by incubation with horseradish peroxidase conjugated secondary antibodies. Immunoreactive bands were visualized with enhanced chemiluminescence Western blotting detection reagents (GE Healthcare, Chalfont St. Giles, UK).
RESULTS
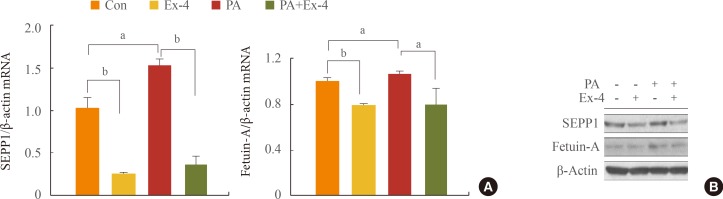
Exendin-4 reduced expression of PA-increased SEPP1 and fetuin-A in human hepatocytes
The expression of the SEPP1 and fetuin-A genes was higher in cells treated with PA alone than in the untreated controls, and significantly decreased with exendin-4 treatment in cells that did and did not undergo PA treatment (Fig. 1A). In addition, the expression of proteins exhibited a trend similar to that of their respective protein transcripts (Fig. 1B).
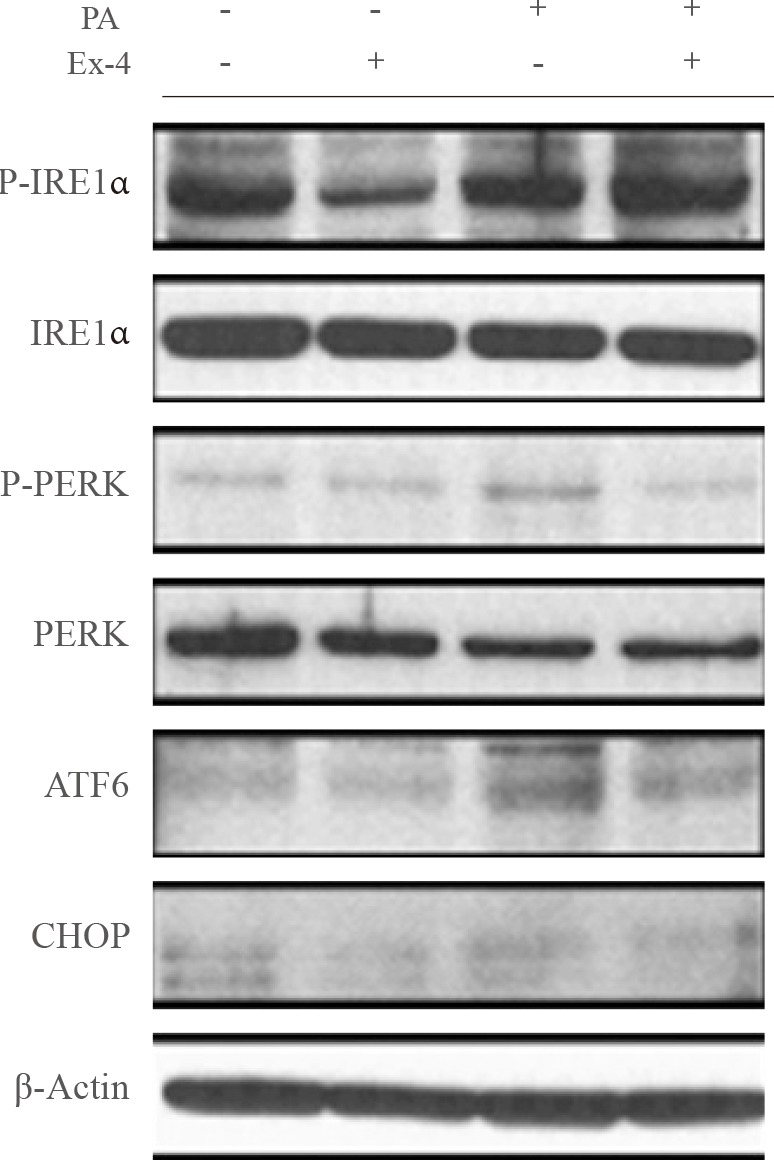
PA, a saturated fatty acid, disrupts ER homeostasis [17], potentially leading to diabetes and hepatic steatosis. To evaluate the effect of exendin-4 on PA-induced ER stress in hepatocyte cells, we examined the effect of exendin-4 on the expression of the ER stress markers IRE1α, PERK, ATF6, and CHOP. Cells exposed to PA displayed higher P-IRE1α, P-PERK, ATF6, and CHOP protein levels, whereas PA-induced increases in the ER stress marker protein levels were reversed in cells treated with exendin-4 (Fig. 2). These results suggest that exendin-4 reduces the expression of PA-induced increases in SEPP1 and fetuin-A, and that exendin-4 improves PA-induced ER stress.
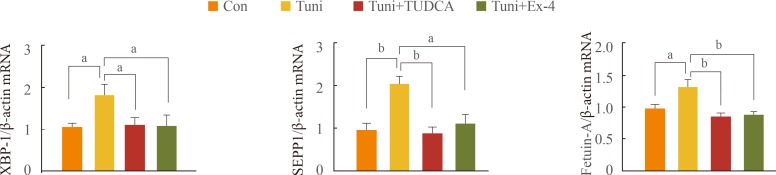
Exendin-4 reduces expression of Tuni-induced SEPP1 and fetuin-A in human hepatocytes
To examine whether increased expression of SEPP1 and fetuin-A secondary to PA treatment is associated with ER stress, HepG2 cells were pretreated with Tuni, an ER stress inducer, followed by the addition of TUDCA, an ER stress inhibitor, or exendin-4. As shown in Fig. 3, the expression of SEPP1 and fetuin-A mRNA as well as XBP-1, a marker for ER stress, was significantly higher in cells treated with Tuni than in the untreated controls. In contrast, supplementation of TUDCA in cells exposed to Tuni completely abolished the effect of Tuni on the expression of these genes. Interestingly, the expression of XBP-1, SEPP1, and fetuin-A in cells treated with exendin-4 exhibited levels similar to those in cells treated with TUDCA. These data suggest that exendin-4 has a protective effect against ER stress, and that exendin-4 attenuates the expression of the SEPP1 and fetuin-A genes by relieving ER stress.
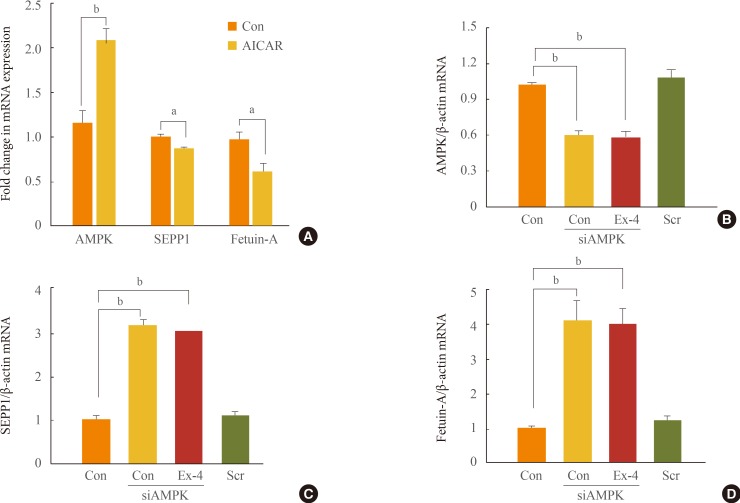
Effect of exendin-4 on regulation of SEPP1 and fetuin-A is mediated via increased AMPK
The AMPK activator AICAR can inhibit fatty acid-induced ER stress [18]. SIRT1-AMPK signaling induces a potent pro-tective effect of exendin-4 against fatty liver disease [19]. In this study, we have demonstrated that AICAR downregulates the expression of the SEPP1 and fetuin-A genes (Fig. 4A). We also examined whether the inhibitory effect of exendin-4 on the expression of ER stress-induced SEPP1 and fetuin-A is mediated by AMPK. The expression of AMPK increased in HepG2 cells treated with exendin-4 [19]. However, when the expression of AMPK was inhibited by AMPK siRNA, the expression of SEPP1 and fetuin-A in cells treated with exendin-4 did not decrease (Fig. 4B-D). These results suggest that exen-din-4 inhibits expression of ER stress-induced SEPP1 and fetuin-A via stimulation of AMPK. Activation of AMPK may mediate an inhibitory effect of exendin-4 on ER stress-induced hepatokines, such as SEPP1 and fetuin-A.
DISCUSSION
We have demonstrated that the expression of SEPP1 and fetuin-A is significantly higher in hepatocytes treated with PA, which induces upregulation of ER stress. On the other hand, exendin-4 treatment decreases the expression of these genes via improvement of ER stress by increasing AMPK.
Recent studies have reported that the hepatokines SEPP1 and fetuin-A can be therapeutic targets of type 2 diabetes mellitus and NAFLD [352021]. Serum SEPP1 and fetuin-A levels are reportedly associated with metabolic syndrome, which is in turn associated with hypoadiponectinemia [62122]. While patients with obesity and NAFLD show significantly increased SEPP1 and fetuin-A levels [423], SEPP1-knockout mice exhibit improved insulin sensitivity in liver and muscle and attenuated adipocyte hypertrophy [3]. Mao and Teng [24] reported that an increased plasma SEPP1 level is the result, rather than the cause of glucose dysregulation, although further studies are needed. In the present study, we demonstrated that the expression of SEPP1 and fetuin-A increased in hepatocytes treated with PA, which can induce fatty liver disease and hepatic insulin resistance [1925], and decreased in hepatocytes treated with exendin-4, which exhibits antidiabetic actions through the GLP-1 receptor. These results are consistent with those of previous studies showing that SEPP1 and fetuin-A are novel biomarkers for metabolic disorders, including obesity, diabetes, and hepatic steatosis [41026].
ER stress plays a crucial role in obesity, insulin resistance [27], and NAFLD [28]. The ER stress response is triggered by different stimuli such as oxidative stress, hypoxia, enhanced protein synthesis, high levels of glucose and saturated fatty acids, and high levels of ER stress inducers, such as thapsigargin and Tuni [2930]. The ER stress response also stimulates ER stress sensors such as ATF6, phosphorylation of PERK and IRE-1α [31], and ER stress-induced transcription factors, such as CHOP and XBP-1 [3233]. In a previous study, we demonstrated that exendin-4 has a protective effect against PA-induced ER stress in hepatocytes [34]. In cells treated with PA, changes in the expression of the ER stress markers, SEPP1 and fetuin-A showed similar patterns, and exendin-4 treatment in cells pretreated with PA or Tuni significantly reduced the expression of these genes. In addition, relieving ER stress by exendin-4 induced decreased expression of the SEPP1 and fetuin-A genes. Thus, these results suggest that decreased expression of SEPP1 and fetuin-A in cells treated with exendin-4 may be associated with improvement of ER stress by exendin-4.
SEPP1 expression is reportedly stimulated by the interaction between the transcription factors FoxO1 and HNF4α, and the coactivator PGC-1α, which stimulates hepatic gluconeogenic gene expression. Jung et al. [35] reported that adiponectin suppresses hepatic SEPP1 through AMPK-mediated phosphorylation of FoxO1a, and ameliorates hepatic fetuin-A through AMPK-mediated reduction of nuclear factor-κB activity [36]. Exendin-4 upregulated the expression of adiponectin and adiponectin receptor 2, and improved hepatic steatosis via SIRT1/AMPK signaling in mice models of diet-induced obesity [19]. The current study suggests that SEPP1 and fetuin-A decreased upon treatment with exendin-4 and the AMPK activator AICAR, whereas when the AMPK gene was silenced with specific siRNA, the SEPP1 and fetuin-A expression did not decrease with exendin-4 treatment. These data suggest that the effect of exendin-4 on the regulation of hepatic SEPP1 and fetuin-A may be mediated by AMPK.
However, the regulatory mechanism of AMPK action on the expression of the SEPP1 and fetuin-A genes remains unclear. Unlike the contention by Jung et al. [35] that salsalate- and adiponectin-mediated AMPK suppresses FoxO1 activity, which is positively associated with SEPP1, Takayama et al. [37] reported that a decrease in the SEPP1 level by treatment with the AMPK activator metformin is dependent on FoxO3a, but not on FoxO1. Moreover, the mechanism of AMPK action on the regulation of fetuin-A expression is poorly defined. Further studies are needed to clarify the effect of exendin-4 on the regulation of hepatic SEPP1 and fetuin-A by treatment with AMPK.
In conclusion, this study demonstrated that PA-induced ER stress stimulates the novel hepatokines SEPP1 and fetuin-A. This study also showed that exendin-4 can suppress the expression of hepatic SEPP1 and fetuin-A via improvement of ER stress by AMPK. Further studies are needed to clarify the details of the mechanism by which exendin-4-induced AMPK controls SEPP1 and fetuin-A.
 XML Download
XML Download