

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Alzheimer's disease (AD) is the most common form of dementia and is a neurodegenerative disorder characterized by the degeneration of neurons, as well as by the progressive decline of cognitive function. AD exhibits the hallmarks of both senile plaques derived from amyloid beta (Aβ) and neurofibrillary tangles, especially in the hippocampus or cerebral cortex, relevant to learning and memory.1,2 When Aβ exists in high concentrations, it forms insoluble and fibrillar Aβ plaques, which activate ion channels in the cell membrane to induce membrane depolarization and destabilization of intracellular calcium homeostasis.3,4,5 In particular, Aβ oligomers cause intracellular Ca2+ overload, leading to neuronal death, which can be prevented by N-methyl-D-aspartate (NMDA) receptor antagonists.6,7
Streptozotocin (STZ) is a glucosamine-nitrosourea compound that, when metabolized, generates a cytotoxic product that preferentially destroys pancreatic β cells.8 The alkylating properties of STZ metabolites generate reactive oxygen species and cause oxidative stress.8 Previously, intracerebroventricular streptozotocin (STZ-icv) administration was shown to induce oxidative stress,9,10,11 neuronal cell damage,12,13 and dysfunctions in learning and memory.8,14,15 Accordingly, STZ-icv models have been used to assess the therapeutic potential of various drugs, as well as other non-drug therapeutic strategies.16 Additionally, STZ-induced learning and memory dysfunction is associated with oxidative stress in animal models.17 Therefore, to attenuate the death of neuronal cells caused by a variety of neurodegenerative diseases, antioxidants have been spotlighted in potential treatments of neurodegenerative diseases such as AD.17,18
Agmatine (Agm) is an endogenous peptide synthesized by arginine decarboxylase, and is reported to be present in glia and neuronal cells. Several researchers have investigated the potential of Agm to improve cognitive function and neuronal cell death in various animal models.19,20,21,22,23 Agm, a neurotransmitter or neuromodulator, exerts neuroprotective effects in various central nervous system injury models, including neurotrauma and neonatal ischemia animal models.24,25,26,27,28 Agm, as an NMDA receptor antagonist, plays a crucial role in regulating the production of nitric oxide (NO). Since NO can enhance a cell's survival rate under oxidative stress, Agm also could protect against damage to cells under oxidative stress.29,30 As well, nuclear factor-erythroid 2 related factor 2 (Nrf2) protects the cell against various stresses and regulates the expression of antioxidant genes, including superoxide dismutase, NAD(P)H, and γ-GCS.31,32,33,34,35,36 Previous studies demonstrated that Nrf2 is related to cognitive decline.37,38,39 Accordingly, this study attempted to investigate whether Agm promotes Nrf2 mediated antioxidant signaling. In particular, we aimed to determine the potential benefits of Agm in improving cognitive dysfunction and preventing cell death in a STZ-induced Alzheimer rat model.
MATERIALS AND METHODS
Animal model
Male Sprague-Dawley rats (n=50, weighing 250-330 g) were used in this study. Rats were maintained under controlled hygienic conditions with a 12 hr light/dark reverse cycle at a constant temperature with free access to food and water. All animal experiments were performed in accordance with the Korean Food and Drug Administration guidelines. Protocols were reviewed and approved by the Institutional Animal Care and Use Committee of the Yonsei Laboratory Animal Research Center (Permit #: 10-115).
Drug treatments
For each experiment, rats were given anesthesia [chloral hydrate 300 mg/kg, intraperitoneal (i.p)], followed by a single injection of STZ (1.5 mg/kg, dissolved in a vehicle consisting of 0.05 M citrate buffer) bilaterally into each lateral ventricle (5 µL/ventricle). Control animals were given an equal volume of intracerebroventricular (icv) vehicle via the same procedure. Agm was purchased from Sigma (Sigma, St. Louis, MO, USA), dissolved in normal saline (pH 7.4), and administered to rats via an intra-peritoneal route. Rats were injected with Agm (100 mg/kg) daily for up to two weeks after surgery. The concentration of Agm (100 mg/kg) was determined based upon the results of our previous study.25
Experimental design
The sham group (n=15) received bilateral icv injection of saline, 5 µL in each rat. The rats in the EC group (n=17) were given injection of STZ-icv (1.5 mg/kg, 1st and 3rd days after surgery) (Sigma, St. Louis, MO, USA) bilaterally.40 Rats in the STZ-Agm group (n=18) were given 5 uL injection of STZ-icv (1.5 mg/kg, 1st and 3rd days after surgery) (Sigma, St. Louis, MO, USA) bilaterally and treated with Agm [100 mg/kg, intraperitoneal (i.p.) daily for two weeks from the 1st day after surgery]. The heads of the rats were positioned in a stereotactic frame (coordinates of 1.5 mm posterior to the bregma, 1.5 mm lateral to the sagittal suture, 2.5 mm ventral to the surface of the brain).
Cognition assessment tests
Cognitive evaluation of rats was tested using a Morris water maze.41 Before conducting the Morris water maze test, we conducted pre-training. For all trials, a single rat was placed in a pool, facing the wall, at a different starting point (NW, NE, SW, or SE), and was then allowed to swim for a maximum of 60 seconds or until it reached the platform. If the rat failed to find the platform during the trial, it was manually guided to the platform by the investigator and placed on top of it for 15 seconds. Next, an apparatus consisting of a circular pool (200 cm diameter, 60 cm high) filled with water (depth 30 cm; 24±1℃) was placed in a room with consistently positioned spatial cues. An escape platform (15 cm diameter) was placed in the middle of one of the quadrants, 1.5 cm below the water surface, equidistant from the sidewall and the middle of the pool. The time required to reach the platform (escape latency) was measured in each trial. After the acquisition stage, a probe test was performed after removing the platform. We measured the latency up to the point when the rat traversed the location where the platform used to be as a measure of memory.
Hematoxylin and eosin (H&E) staining
Rats were anesthetized and perfused as previously mentioned.42 Rat brains were cut into coronal slices of 2 mm thickness using a rat brain matrix (Ted Pella, Redding, CA, USA). Next, the brain slices were fixed with 4% paraformaldehyde (pH 7.4) for 3 days and subsequently embedded in a paraffin block. The paraffin blocks containing the hippocampus and cortex were deparaffinized and re-hydrated with different concentrations of alcohol and xylene. After hematoxylin and eosin (H&E) staining, stained hippocampus and cortex sections were examined under a microscope.
Immunohistochemistry
Five µm thick frozen brain sections were harvested onto clean glass slides (Thermo Scientific, Bremen, Germany), air-dried, and fixed in cold acetone for 10 minutes at -20℃. The slides were washed first in Tris-buffered saline (TBS) and then incubated with 0.3% H2O2 in methanol to quench endogenous peroxidase activity. Followed by a series of washes (three times with distilled water), the sections were blocked with 10% normal rabbit serum. Frozen brain sections (20 µm) were fixed in ice-cold acetone for 20 minutes. To block specific staining, sections were incubated in 5% bovine serum albumin (BSA) (Sigma-Aldrich, Sigma, St. Louis, MO, USA) diluted in PBS for 30 minutes before addition of primary and secondary antibodies. Primary antibodies for phosphorylated insulin receptor substrate-1 (p-IRS-1) (1:200, Santa Cruz, CA, USA), 8-oxo-2'-deoxy-guanosine (8-OHdG) (1:200, Chemicon, Billerica, MA, USA), and anti-Aβ (1:200, Millipore, Billerica, MA, USA) were applied to the samples for 24 hours at 4℃, followed by a 90-minute incubation with appropriate florescence secondary antibody (1:100, Invitrogen, Carlsbad, CA, USA) and three washings in PBS for 10 minutes each. After three washes in 0.1% phosphate buffered saline with Tween-20 (PBST), the sections were incubated with rhodamine-conjugated sheep anti-rabbit or sheep anti-mouse secondary antibody that was diluted to 1:200 with 5% BSA fraction V in 0.1% PBST for 2 h in the dark at room temperature. After three washing in PBS, all sections were incubated with 1 µg/mL of 4',6-diamidino-2-phenylindole (Sigma-Aldrich, Sigma, St. Louis, MO, USA) and 2 µg/mL of propidium iodide (Sigma-Aldrich, USA) for a counter staining. Tissues were then visualized under a confocal microscope (Zeiss LSM 700, Carl Zeiss, Thornwood, NY, USA).
Western blot analysis
For Western blot analysis, rats from all groups were sacrificed and their brains were perfused with saline through the heart aorta to rinse away traces of blood. Next, the portion of the brain comprising the hippocampus and cortex was dissected for extraction of proteins and treated with lysis buffer (1% Triton X-100, 0.5% NP-40, 150 mM NaCl, 10 mM Tris (pH 7.4), 1 mM Ethylene-di-amine tetra acetic acid, 1 mM ethylene glycol tetraacetic acid (pH 8.0), 0.2 mM sodium orthovanadate, 0.2 mM phenyl methyl sulfonyl fluoride, and protease inhibitor cocktail). Isolated proteins were centrifuged at 12000 rpm at 4℃. Equal amounts of protein (20 µg) from the supernatants were separated on a 10% acrylamide gel and proteins were electrophoretically transferred onto nitrocellulose membranes. After blocking with 5% skim milk for 2 hrs, the membranes were incubated with primary antibodies against Bcl2 (1:1000, Santa Cruz, San Jose, CA, USA), Bax (1:1000, Santa Cruz, San Jose, CA, USA), cleaved caspase-3 (1:1000, Cell signaling, Billerica, MA, USA), PI3K (1:2000, Millipore, Billerica, MA, USA), Nrf2 (1:200, Santa Cruz, San Jose, CA, USA), γ-GCS (1:500, Thermo Scientific, Bremen, Germany), and β-actin (1:1000, Millipore, Billerica, MA, USA) at 4℃ overnight. Later the membranes were washed three times for 5 min each with TBST. The detection of secondary anti-rabbit and anti-mouse antibodies (1:3000, New England Bio labs, USA) was conducted for 1 hour at room temperature. After washing with PBST (0.05% with Tween 20) three times, immunoreactive signals were detected by chemiluminescence with an ECL detection system (Amersham Life Science, London, UK) using the LAS 4000 program.
Statistical analysis
Statistical analyses were carried out using SPSS 18.0 software (IBM Portsmouth, IBM North Harbour, Portsmouth, UK). All data are expressed as means±S.E.M. Statistical significance in intergroup differences was determined by one-way analysis of variance, followed by Scheffe's post hoc multiple comparison test. Each experiment included at least three replicates per condition. Differences with a p value less than 0.05 were considered statistically significant.
RESULTS
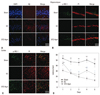
Agmatine treatment attenuates Aβ accumulation and improves cognitive dysfunction in STZ-icv rats
To determine the accumulation of Aβ, on the 21st day following STZ injection, Aβ staining was performed in the sham, EC, and STZ-Agm groups. The expression of Aβ was considerably decreased in the STZ-Agm group compared to the EC group (Fig. 1A). To confirm phosphorylation of IRS-1 by Agm treatment in STZ-icv rats, we conducted immunohistochemistry using phospho-IRS-1 antibody, because IRS-1 plays a key role in transmitting signals from the insulin receptors to intracellular pathways. In the STZ-Agm group, both hippocampus (Fig. 1B) and cortex regions (Fig. 1C) showed an increase in IRS-1 phosphorylation compared to the EC group. These data indicated that Agm treatment could promote IRS-1 phosphorylation in STZ-icv rat models.
In previous studies, Morris water maze tests have been commonly applied for the assessment of cognition and memory functions. To confirm the enhancement of memory function in our STZ-induced Alzheimer rat model upon treatment with Agm, we conducted Morris water maze tests. The swimming times of four trials per day for 5 days in each group are shown in Fig. 1. Escape latency time (days 1-5) (to find a hidden platform) was significantly prolonged in the EC group compared to the sham group (Fig. 1D). Fig. 1D shows that the STZ-Agm group animals presented a significantly lower latency to find the platform than the EC group. The animals of the STZ-Agm group demonstrated improved Morris water maze acquisition performance. Our findings suggest that Agm treatment could improve learning and memory dysfunction in STZ-induced Alzheimer rats.
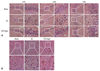
Agmatine treatment attenuates histological abnormalities in hippocampus and cortex regions of STZ-icv rats
To investigate morphological differences in cells, we conducted H&E staining. The H&E staining of Cornu Ammonis (CA)1, CA2, and CA3 regions of the hippocampus revealed a large number of degenerated cells in the EC group compared with STZ-Agm group (Fig. 2A), which were evidenced by a decrease in the number of H&E stained cells. The EC group exhibited greater shrinkage of the nuclei of cells than those in the STZ-Agm group. Fig. 2B shows that the number of abnormal cells in the cortical region were higher in the EC group than the STZ-Agm group. Fig. 2 indicates that Agm treatment reduces STZ-induced histological abnormalities in hippocampus and cortex regions, compared with the EC group.
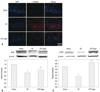
Agmatine treatment inhibits the cell death pathway in STZ-icv rats
To confirm apoptotic cell death in the EC group and the STZ-Agm group, western blot analysis was conducted to check expression of apoptotic proteins. Fig. 3A indicates that the presence of cleaved caspase-3, an active form of caspase-3, was attenuated in the STZ-Agm group compared with the EC group. Additionally, in the present study, the anti-apoptotic effect of Agm was investigated by checking the expression of proteins, such as Bax, Bcl2, and PI3K, using western blotting. Among the quantitative western blot results, the expression of Bax, known as a pro-apoptotic protein, was higher in the EC group than the STZ-Agm group (Fig. 3C). In contrast, Bcl2, known as an anti-apoptotic protein, was expressed more in the STZ-Agm group than the EC group (Fig. 3B). Also, the STZ-Agm group showed increased expression of PI3K, known to be related to survival pathways and insulin signal transduction, compared with the EC group (Fig. 3D). Fig. 3 suggests that Agm treatment inhibits cell death signaling in STZ-induced Alzheimer rats.
Agmatine treatment promotes the Nrf2-mediated antioxidant pathway in STZ-icv rats
To detect the generation of Reactive Oxygen Species (ROS), which causes DNA damage to the cells, we conducted 8-OHdG staining. The immunohistochemical localization of 8-OHdG revealed a decreased number of 8-OHdG positive cells in the STZ-Agm group compared to the EC group (Fig. 4A). Fig. 4A indicates that ROS generation is decreased in the STZ-Agm group compared with the EC group. The western blot data for Nrf2 (known as an antioxidant transcription factor) (Fig. 4B) and γ-GCS (an important enzyme in glutathione synthesis) (Fig. 4C) show that Agm treatment promotes Nrf2-mediated antioxidant pathway signaling. Fig. 4 indicates that Agm treatment prevents cell death and promotes Nrf2-mediated antioxidant signaling in STZ-induced Alzheimer rats.
DISCUSSION
AD is one of the most prevalent degenerative diseases in the elderly. AD is characterized by progressive memory loss and cognitive impairment.43 The administration of STZ to the central nervous system (CNS) generates similar pathology to AD.44,45,46 Previously, STZ-icv administration was shown to induce oxidative stress, neuronal cell damage, and dysfunction in learning and memory.8,14,15 Accordingly, STZ-icv models have been used to assess the therapeutic potential of various drugs. Hence, we employed a STZ-induced Alzheimer rat model to investigate AD like pathologies.
In previous studies, the administration of STZ in the CNS generated similar pathology to AD, mainly accumulation of Aβ.46,47 In regards to the pathologic status of AD, the hyper activation of glutamate receptor and continuous Ca2+ influx by Aβ results in neuronal damage and cognitive dysfunction.48,49,50 Additionally, a previous study showed that memantine, as a glutamate NMDA receptor channel antagonist, blocks NMDA overstimulation upon excitoxicity; accordingly, memantine was suggested to be of potential use in the treatment of AD.51 Several studies reported the neuroprotective effects of Agm in a variety of neurodegenerative pathologies through possible blockade via NMDA channels.24,25,30,52,53,54 As shown in our data, Aβ accumulation in damaged brain regions was decreased in the STZ-Agm group compared to the EC group.
Also, STZ-icv in rats can induce brain insulin system dysfunction and induce progressive deficits in learning, memory, and cognitive behavior like sporadic AD.12,46 As shown in our data, STZ inhibits phosphorylation of IRS-1 in rats. IRS-1 plays a key role in transmitting signals from insulin receptors to intracellular pathways.55 Accordingly, several studies demonstrated that IRS-1 signaling pathways are associated with cognitive function in CNS diseases.56 Insulin dysregulation contributes to AD pathologies by reducing brain glucose utilization.57,58,59 Insulin affects neuronal cognition and memory by regulating ion channels and neurotransmitter receptors in AD brains.60,61 Considering that insulin signal transduction is important for cognitive function,59,62,63,64 our data indicated that Agm could induce the activation of brain insulin signal transduction and improve learning and cognitive decline in STZ-induced Alzheimer rat model.
In addition, Agm inhibits cell death by regulating the production of NO,29,30,65 and also attenuates neuronal cell death in neurodegenerative animal models.19,20,21,22,23,66,67,68 During the onset of AD, there is increased oxidative stress leading to the retardation of cognitive ability.69,70,71 In the present study, water maze test results highlighted significant improvements in test scores in the STZ-Agm group, compared with EC group. These functional outcomes may because Agm, a NMDA antagonist, improves memory function in EC rats.72,73 Additionally, our H&E staining data indicated that the excessive production and accumulation of ROS by Aβ can cause functional and structural changes in critical macromolecules leading to lipid peroxidation, protein oxidation, and DNA cleavage. Earlier studies have suggested that antioxidant treatment could therapeutically cure and prevent neurodegenerative diseases, especially sporadic AD.17,18 Moreover, previous researchers reported that antioxidants such as melatonin, vitamin E, and selegiline can be used to cure AD.74,75,76,77
Also, STZ-induced learning and memory dysfunction is considerably associated with oxidative stress in animal models.78,79,80 The levels of molecular markers for DNA (particularly 8-OHdG) are reported to be elevated in the brains of patients with AD.72 Bcl2 is neuroprotective against apoptotic cell death caused by Aβ.81 Accordingly, overexpression of Bcl2 could attenuate the processing of amyloid precursor protein and tau and reduce extracellular deposits of Aβ.82 Bcl2 protects neuronal cells by inhibiting the activation of caspase-3.81,82 Previously, Bcl-2 expression was shown to be upregulated, while Bax and caspase-3 were down regulated in AD models.83 Bax also plays an essential role in oligomeric Aβ-induced neuronal cell death.84 In the present study, Bax and Bcl285,86 were measured. Agm treatment significantly decreased the expression of 8-OHdG, cleaved caspase-3, and Bax, and increased the expression of Bcl2 in STZ-induced Alzheimer rats.
PI3K signaling cascade promotes NO generation through the activation of endothelial nitric oxide synthase.63,87 In addition, the PI3K/Akt signaling pathway plays crucial roles in cell survival, growth, gene expression, apoptosis, metabolism,88 and also, neuronal survival.89 Liu, et al.64 demonstrated that the insulin-PI3K/Akt signaling pathway is reduced in AD brain. As IRS-1 plays a key role in transmitting signals from the insulin receptors to intracellular pathways including the PI3K pathway, the phosphorylation of IRS-1 by Agm treatment could improve cognitive decline and protect against cell death by activating the PI3K pathway.
Nrf2 plays an important role in regulating cellular oxidative stress and controls the expression of many detoxifying genes such as catalase, superoxide dismutase, heme oxygenase-1 (HO-1), NAD(P)H, and γ-GCS.31,32,33,34 Nrf2 activation can induce the antioxidant pathway and protect cells against oxidative stress.35,36 In addition, Nrf2 ameliorates cognitive impairment.37,38,39 Our western blot data for Nrf2 and γ-GCS indicated that Agm could promote Nrf2-mediated antioxidant pathways in STZ-induced Alzheimer rats.
In conclusion, Agm could improve cognitive decline by decreasing accumulation of Aβ and ameliorating insulin signal transduction. Also, Agm could protect against damage to cells by activating Nrf2-mediated antioxidant signaling. Hence, Agm may be a promising therapeutic agent for alleviating neuronal cell apoptosis and cognitive decline in AD.

 XML Download
XML Download