

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetic nephropathy (DN), a chronic microvascular complication of diabetes mellitus, is one of leading causes of end-stage renal disease (ESRD) in industrialized countries.1 Emerging evidence suggest that DN leads to tubular injury caused by tubular atrophy, as reflected by increased apoptosis in proximal and distal tubular epithelia in patients with diabetes.2 Human Na+/H+ exchanger isoform 1 (NHE1) is a widely expressed membrane protein that plays a critical role in maintaining intracellular pH (pHi) balance.3 It has been demonstrated that cellular NHE1 activity is elevated in type 1 patients with nephropathy and essential hypertension.4 The increased NHE1 may disturb the inner acid-base balance and contribute to progressive renal impairment. Meanwhile, emerging evidence showed that NHE1 is involved in high glucose (HG) or hyperglycemia induced vascular injury and is associated with the development of DN.5 However, the role of NHE1 in HG induced apoptosis of proximal tubule cell and the underlying mechanism remains poorly understood.
NHE1 is an 815 amino acid glycoprotein that functions by extruding intracellular protons in exchange for extracellular sodium. The NHE1 protein is composed of a 14 amino acid cytoplasmic tail, a 12-helical transmembrane domain responsible for Na+/H+ exchange, and a 315 amino acid cytoplasmic domain playing a regulatory role.6 In addition, NHE also functions in cellular volume regulation and as a scaffolding platform for the assembly of signaling complexes.7 NHE activity is minimal under basal conditions, while it is activated by multiple extracellular stimuli, such as hormones, shear stress, and other pathological factors, which in turn lead to the change of many cell behaviors, including cell proliferation, apoptosis, and migration.8 Moreover, patients with DN manifest with disturbances in electrolytes and the acid-base balance caused by irregular glucose metabolism.910 Therefore, targeting the abnormal NHE1 in tubule cells under diabetic stress might rescue damaged kidney and disturbed electrolytes metabolism.
Our study aimed investigate the impacts of HG on NHE expression and activity, the effects of NHE modulation on HGinduced proximal tubule cell apoptosis, and the underlying mechanism of HG-induced NHE1 dysfunction to highlight a potential therapeutic strategy for the protection of DN.
MATERIALS AND METHODS
Cell culture
Human papillomavirus 16 (HPV-16) transformation immortalized human proximal tubule cell line HK-2 was obtained from American Type Culture Collection, and was grown in DMEMF12 culture medium (GIBCO, Invitrogen, Grand Island, NY, USA) supplemented with 10% FBS (GIBCO, Invitrogen) and 5 mmol/L glucose at 37℃ in a 5% CO2 incubator. For examination of the effects of HG on cellular activity involving NHE1, the HK-2 cells were plated at a density of 5×105 cells on 60 mm dishes containing 5 mM glucose. The serum was withdrawn for 24 h when the cells reached 90% confluence, and they were then treated with HG (25 mM) alone or pretreated with 20 nM NHE1 inhibitor 5-(N,N-dimethyl)-amiloride (DMA) or 20 nM Cyclic GMP-Dependent Protein Kinase Type I (PKG) inhibitor KT-5823 (Sigma, St. Louis, MO, USA) for 2 h under serum-free conditions before exposure to HG. The incubated cells were collected for the following experiments.
NHE1 siRNA transfection
HK-2 cells were seed in a six-well plate at a density of 2×105 cells per well with 2 mL antibiotic-free DMEM-F12 medium supplemented with FBS, and cultured at 37℃ in a CO2 incubator until the cells were 60–80% confluent. A small siRNA duplex against NHE1 (6 µL) and the corresponding negative control siRNA was diluted in 100 µL transfection medium and then mixed with diluted Lipofectamine 2000 (Invitrogen) according to the manufacturer's protocol. For each transfection, siRNA transfection mixture was added to the washed cells, and cells were incubated for 8 h at 37℃ in a CO2 incubator. After transfection, cells were allowed to grow under regular conditions for 48 h. The efficiency of NHE1 silencing was measured using a Western blot assay.
Western blot
Expression of NHE1 and PKG in HK-2 cells was analyzed using a Western blot with GAPDH as an internal control. After being treated with HG alone or in combination with NHE1/PKG inhibitor, the cells were lysed with RIPA and the whole lysates were then subjected to SDS-PAGE. The separated proteins were subsequently transferred to a PVDF membrane using a transfer cell system (Bio-Rad, California, CA, USA). After blocking with 5% milk powder for 1 h, the membrane was incubated with primary antibodies against NHE1 (Santa Cruz, CA, USA), PKG (StressGen Bioreagents, Victoria, BC, Canada) and GAPDH (CST, Danvers, MA,USA) overnight at 4℃. After being washed three times, the membrane was incubated with HRP-labeled secondary antibodies. Blot signals were visualized using Amersham western blot detection reagents (GE Healthcare, Piscataway, NJ, USA). Band densitometry quantification was performed using ImageJ software (ver. 1.43u; National Institute of Health, Bethesda, MD, USA).
Cell apoptosis
Cell apoptosis was determined using Annexin V-FITC and propidium iodide (PI) double-labeling kits according to the manufacturer's recommendations (BD Biosciences, San Jose, CA, USA). The stained cells were observed in a FACSAria III cytometer (Becton Dickinson, San Jose, CA, USA). A total of at least 20000 events were analyzed per sample. Data were analyzed using FACSDiva software (Becton Dickinson).
Reactive oxygen species determination
Reactive oxygen species (ROS) production in HK-2 cells in response to HG treatment was detected as described previously.11 The HK-2 cells were plated on six-well plates at 5×104 cells per well, cultured overnight, and then pretreated with or without NHE1/PKG inhibitor before being incubated with 30 mM HG. DMSO and N-(4-hydroxyphenyl) retinamide (4HPR) (Sigma-Aldrich, St. Louis, MO, USA) were used as controls. After 48 h of incubation, the cells were harvested, washed in PBS, and then loaded with 50 µM 2',7'-dichlorofluorescein-diacetate (DCFH-DA) at 37℃ for 10 min. Fluorescence was detected using a FACSCanto cytometer (BD) on the FL1 channel and analyzed with FACSDiva 6.0 software.
8-OHdG staining
8-hydroxy-2'-deoxyguanosine (8-OHdG) staining was performed to indicate DNA damage in HK-2 cells by HG involving NHE1 regulation. The treated cells were incubated with a mouse monoclonal antibody against 8-OHdG (Abcam, Cambridge, UK) for 8 h, and then a rabbit anti-mouse IgG-FITC was added. Cell nuclei were visualized by 1 mg/mL DAPI (Sigma, St. Louis, MO, USA) staining for 1 min. Cells were then observed under an Olympus fluorescence microscope.
pHi determination
pHi was determined using a pHi-sensitive probe, as previously described.12 In brief, HK-2 cells and/or NHE siRNA transfected HK-2 cells were treated with or without HG for 48 h. Afterwards, the cells were plated on coverslips and incubated with 2 mM DMSO-dissolved pH probe 2',7'-bis-(2-carboxyethyl)-5-(and -6)-carboxyfluorescein (BCECF AM) (AAT Bioquest, Sunnyvale, CA, USA) at 37℃ for 30 min. Then, the coverslips were placed in an open bath imaging chamber containing HCO3-free HEPES-MEM, and the chamber was fixed on an Olympus inverted epifluorescence microscope. The fluorescence emissions at 535 nm were recorded. The pHi was estimated according to the standard curve of the manufacturer's instructions.
Statistical analysis
All statistical analyses were performed using SPSS17.0 software (SPSS Inc., Chicago, IL, USA), and all data are presented as means±SEM. Statistical comparisons were performed using one-way analysis of variance followed by the Student-Newman-Keuls test. All p-values less than 0.05 were considered to indicate statistical significance.
RESULTS
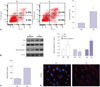
High glucose induces apoptosis and oxidative stress in HK-2 cells
In order to observe the injury effects of HG on renal proximal tubule cells, we first determined changes in apoptosis and oxidative stress in HK-2 cells in response to HG stimulation. As shown in Fig. 1A, the cell apoptosis proportion was 9.21±2.04% under regular conditions with exposure to 5 mM glucose, while cell apoptosis significantly increased to 30.51±6.35% after 30 mM glucose stimulation at 48 h (p<0.05). In addition, proapoptotic proteins, such as Bax and Caspase 3, significantly increased in HK-2 cells following HG insults and were accompanied by a decrease in anti-apoptotic Bcl-2 (Fig. 1B). Furthermore, we evaluated oxidative stress by determining the production of ROS and staining of 8-OHdG. Therein, HG dramatically induced the production of ROS and 8-OHdG (p<0.01) (Fig. 1C and D).
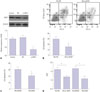
High glucose upregulates NHE1 expression and pHi
Since NHE1 is of importance in maintaining pHi and cell growth, we intended to evaluate its expression and function in HK-2 cells in response to insults by HG. In doing so, we found that HG upregulated the expression of NHE1 in a time-dependent manner, and the NHE1 significantly increased after 24 h as determined by western blot assays (p<0.05) (Fig. 2A). Meanwhile, pHi significantly increased to 7.5 after HG treatment at 24 h, while there was no marked change of pHi in HK-2 cells incubated in normal glucose (Fig. 2B).
NHE inhibition represses high glucose-induced HK-2 apoptosis
In order to investigate the role of NHE1 in HG-induced nephropathy, NHE1 siRNA was introduced to HK-2 cells to suppress its expression (Fig. 3A). NHE1 siRNA transfection significantly reduced cell apoptosis, compared to that transfected with negative control siRNA (p<0.05) (Fig. 3B). In addition, functional inhibition of NHE1 by its antagonist DMA also suppressed cell apoptosis (Fig. 3C). Moreover, we found that both NHE siRNA and its antagonist DMA reduced [pH]i and promoted the pH hemostasis disturbed by HG (Fig. 3D).
PKG is involved in high glucose-induced NHE1 dysregulation
Studies have shown that oxidative stress is responsible for the regulation of NHE1 expression.1314 Therefore, we observed the effects of HG-created oxidative stress conditions on the expression of NHE1. Treatment of HG-exposed HK-2 with antioxidants, such as glutathione (GSH) and N-acetyl cysteine (NCys), significantly abrogated ROS production, accompanied with a reduction in NHE1 expression. However, ROS and NHE1 expression increased greatly after stimulation with H2O2 in HK-2 cells (Fig. 4A and B). Moreover, we investigated whether PKG is involved in HG-induced ROS production and subsequent NHE1 upregulation. PKG significantly increased in HGincubated HK-2 cells, compared to that cultured in normal glucose (p<0.05) (Fig. 4C). In addition, PKG inhibitor KT-5823 markedly suppressed HG-induced HK-2 cell apoptosis, compared with control cells (Fig. 4D). Meanwhile, exogenous cyclic GMP activated PKG and promoted the production of ROS and the expression of NHE1. However, PKG inhibitor KT-5823 suppressed ROS and the expression of NHE1 (Fig. 4E and F).
DISCUSSION
The prevalence of diabetes is rapidly increasing and is accompanied with a high incidence of the resultant kidney damage, termed as DN, which greatly increases the risk of renal failure. Patients with DN manifest with disturbances in electrolytes and in the acid-base balance, as well as circulating uremia toxins caused by disorder of glucose metabolism or other medications.910 NHE1 is a critical Na+/H+ exchanger and plays an important role in maintaining the balance of pHi. However, its role in the pathogenesis of DN is unclear. Our study demonstrated that HG increases oxidative stress, which is responsible for the induction of NHE1 and proximal tubule cell death, eventually leading to tubular injury.
The kidneys play a critical role in maintaining acid-base balance in humans and other mammals.15 As a ubiquitously expressed isoform of a Na+/H+ exchanger, NHE1 serves as a housekeeping protein in the regulation of pHi via the exchange of extracellular Na+ with H+ in a 1:1 ratio. A previous study showed that NHE1 together with NHE3 are of great importance in regulating pHi in rat inner medullary thin limbs (Henle's loop) via increasing H+ extrusion.16 In this study, we observed that HG induces irregular NHE1 expression and cellular pHi in tubular cells. Meanwhile, however, NHE1 inhibitor restores this imbalance after HG insult. Moreover, pHi homeostasis is also of importance for cell volume, function, and survival. Therefore, NHE1 inhibition under HG stress facilitates the maintenance of pHi homeostasis and subsequent cell survival.
NHE1 is also responsible for the regulation of multiple cellular functions, such as cell motility and cell survival, by its interaction with the cytoskeleton, as well as a variety of cytoplasmic proteins.17 It is well recognized that NHE1 presents on the basolateral plasma membrane of epithelial cells and plays an important role in defending against apoptosis in naïve proximal tubule cells. While the prosurvival effect of activated NHE1 can be surmounted, cells proceed toward apoptosis when they are exposed to overloaded or sustained apoptotic stimulus.18 As reported by Li, et al.,5 advanced glycation end products (AGEs) induce kidney dysfunction accompanied by a marked increase in NHE1. The authors showed that elevated NHE1 broke the membrane integrity of renal cortex, rather than exerting a protective role.5 Moreover, it was also claimed that NHE1 overexpression further aggravates cell apoptosis in cardiomyocytes following a hypoxia/reoxygenation challenge.3 However, the role of NHE1 in HG induced apoptosis of proximal tubule cell remains unclear. We demonstrated that HG stimulates the overexpression of NHE1 and NHE1-mediated pH dysregulation and that inhibition of NHE1 restores the pH disorder and cell apoptosis induced by HG.
Diabetes is a serious metabolic disorder that causes oxidative stress and inflammation, which contribute to its pathogenesis and complications.19 Accumulated evidence has shown that HG or hyperglycemia exacerbates oxidative stress and the resultant epithelial-mesenchymal transition, as well as apoptosis, in renal tubular epithelial cells.202122 Furthermore, we showed that HG-induced ROS are responsible for the induction of NHE1 in renal tubular cells and that NHE1 may participate in HG-induced cell death. It was reported that oxidative stress mediated by H2O2 or superoxide increased NHE1 activity in other cells, such as rat ventricular myocytes and thick ascending limb.1314 In addition, we found that PKG is involved in the induction of oxidative stress and NHE1 activity. Elevating cGMP has been shown to increase the potassium current, which is mediated by the accumulated intracellular ROS in microglia cells.23 However, role of PKG-ROS axis in inducing NHE1 in tubular cells has not been revealed yet. Nevertheless, our study suggests the potential role of NHE1 in HG induced tubular cell apoptosis involving oxidative stress; however, further in vivo studies concerning the role of NHE1 in tubulopathy in DN animals and the effect of the PKG-ROS axis in inducing NHE1 are required.
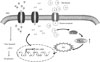
Collectively, our study demonstrated that HG induces higher oxidative stress and cell apoptosis in renal tubular epithelia cells. The PKG signaling triggered excessive oxidative stress that was involved in the regulation of NHE1 expression and functions in response to HG stress (Fig. 5). NHE1 dysregulation may serve as a proapoptotic process in promoting tubular cell death. Our study may provide evidence for understanding metabolic acidosis in the progression of DN. However, more details related to NHE1 in kidney injury are needed.


 XML Download
XML Download